Плазмида pET24bxylA, обеспечивающая синтез ксилозоизомеразы в клетках рекомбинантного штамма Escherichia coli БИМ В-427 Д, и рекомбинантный штамм Escherichia coli БИМ В-427 Д – продуцент ксилозоизомеразы
Номер патента: 12214
Опубликовано: 30.08.2009
Авторы: Сапунова Леонида Ивановна, Лобанок Анатолий Георгиевич, Евтушенков Анатолий Николаевич, Шляхотко Екатерина Александровна







Текст
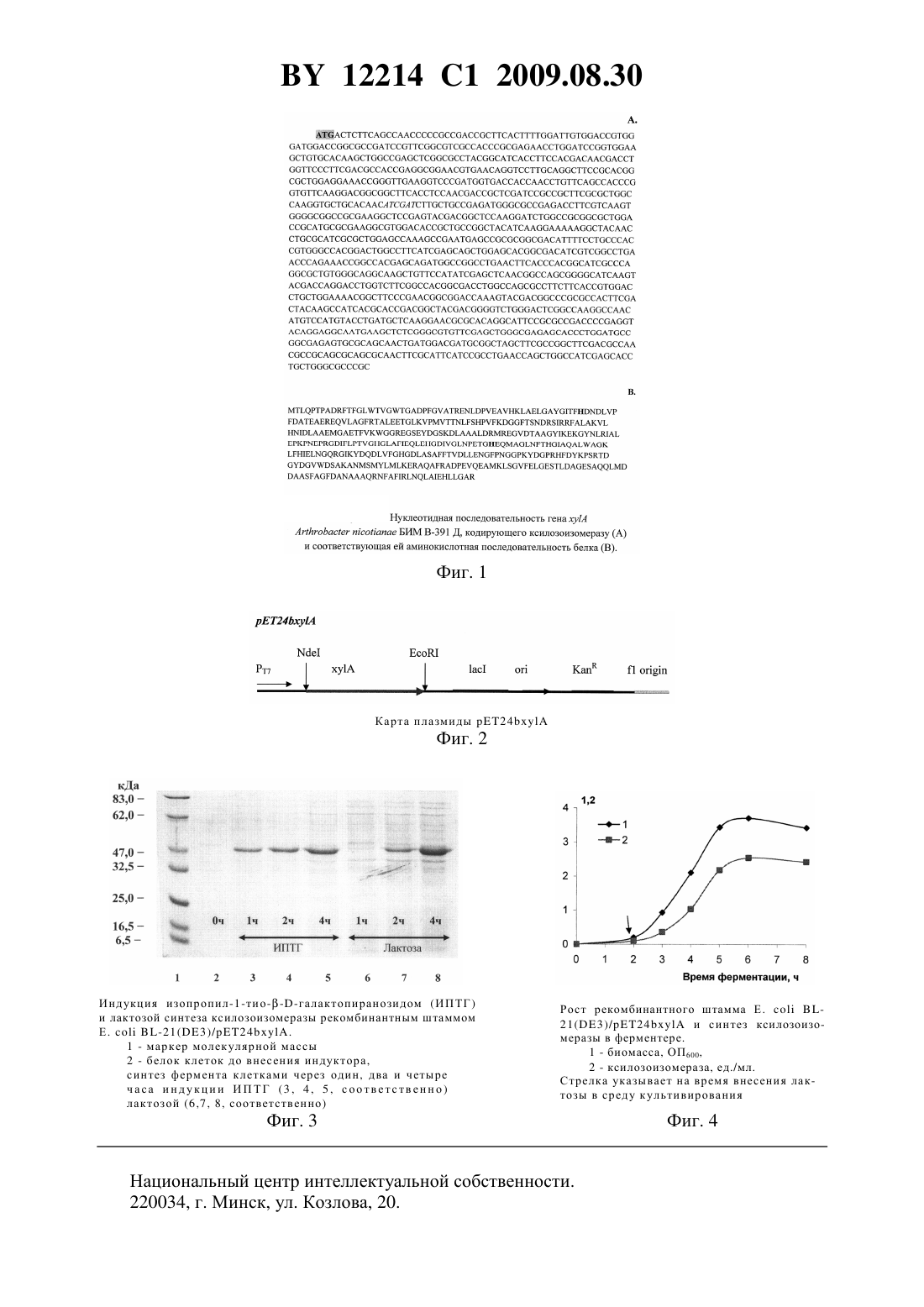
(51) МПК (2006) НАЦИОНАЛЬНЫЙ ЦЕНТР ИНТЕЛЛЕКТУАЛЬНОЙ СОБСТВЕННОСТИ ПЛАЗМИДА 24, ОБЕСПЕЧИВАЮЩАЯ СИНТЕЗ КСИЛОЗОИЗОМЕРАЗЫ В КЛЕТКАХ РЕКОМБИНАНТНОГО ШТАММАБИМ В-427 Д,И РЕКОМБИНАНТНЫЙ ШТАММБИМ В-427 Д - ПРОДУЦЕНТ КСИЛОЗОИЗОМЕРАЗЫ(71) Заявитель Государственное научное учреждение Институт микробиологии Национальной академии наук Беларуси(72) Авторы Шляхотко Екатерина Александровна Сапунова Леонида Ивановна Лобанок Анатолий Георгиевич Евтушенков Анатолий Николаевич(73) Патентообладатель Государственное научное учреждение Институт микробиологии Национальной академии наук Беларуси(57) 1. Плазмида 24, содержащая встроенную по сайтам рестрикцииив область полилинкера плазмиды 24 нуклеотидную последовательность, кодирующую ген ксилозоизомеразыБИМ В-391 Д, которая фланкирована промотором и терминатором фага Т 7, обеспечивающая синтез ксилозоизомеразы в клетках рекомбинантного штамма-21(3)/24 БИМ В-427 Д. 2. Рекомбинантный штамм-21(3)/24 БИМ В-427 Д, содержащий плазмиду по п. 1, продуцирующий ксилозоизомеразу. Изобретение относится к микробиологии и биотехнологии, в частности к генной и белковой инженерии, и может быть использовано для получения новых рекомбинантных штаммов и препаратов ксилозоизомеразы, применяемой в промышленности для производства из осахаренного крахмалсодержащего сырья натуральных подсластителей - глюкозо-фруктозных сиропов и кристаллической фруктозы. Для получения глюкозо-фруктозного сиропа и фруктозы используется микробная ксилозоизомераза, производство которой занимает лидирующие позиции в области биотехнологии ферментных препаратов. Основу рентабельности коммерческого производства продуктов микробного синтеза, включая препараты ксилозоизомеразы, составляют высокоактивные штаммы-продуценты. Главенствующая роль в их создании на современном этапе развития науки принадлежит технологиям рекомбинантных ДНК. Синтез ксилозоизомеразы микроорганизмами обусловливается наличием в их геноме гена . Согласно имеющимся в научно-технической литературе данным, при условии,что клонированный геннаходится под контролем -, -, - и других сильных 12214 1 2009.08.30 промоторов, на долю ксилозоизомеразы, продуцируемой генетически модифицированными штаммами, приходится от 20 до 60 совокупного внутриклеточного белка 1-6. Известен продуцент термоустойчивой ксилозоизомеразы - рекомбинантный штамм, наследующий плазмидную ДНК, в которой ген ксилозоизомеразы из 33223 клонирован под контролем промотора бациллярного гена, ответственного за синтез белка клеточной стенки 7. Новый рекомбинантный штамм характеризовался в 100 раз более высокой, чем исходный штамм 33223, продуктивностью. Однако использованная при конструировании штамма система контроля экспрессии гена не позволяет ему достичь продуктивности, обеспечивающей рентабельность производства ферментного препарата. Известен рекомбинантный штамм-21, у которого ген , кодирующий термостабильную ксилозоизомеразу-1 (-11116), и контролирующий его промотор фага Т 7 локализован на плазмиде 8. Необходимым условием экспрессии гена в клетках рекомбинантного штамма, ограничивающим его широкое использование для коммерческого получения ксилозоизомеразы, является обязательное наличие в питательной среде дорогостоящего индуктора - изопропил-1-тиогалактопиранозида. Известен рекомбинантный штамм, содержащий гибридную плазмиду 15 с геном ксилозоизомеразы 319. Уровень продукции ксилозоизомеразы трансформантом, в отличие от исходного штамма, повышен в 2,7 раза,однако удельная активность фермента составила только 0,148 ед/мг белка 9. Известно существенное повышение уровня синтеза фермента рекомбинантным штаммом 109/12 при клонировании в нем гена ксилозоизомеразы, котролируемого -промотором. Продуктивность нового штамма была в 45 раз более высокой, чем исходного, но при этом составляла лишь 450 М/минл среды 10. Известно, что экспрессия в клетках рекомбинанта-21(3)/10 гена ксилозоизомеразы/- 489, находящегося под контролем промотора фага Т 7/ас, индуцируется изопропил-1-тиогалактопиранозидом, и доля фермента в объеме синтезируемого бактериями белка составляет 1511. Известны также рекомбинантные штаммы-21(3)/рЕТ 23 аТТХ и-21(3)/23, которые несут индуцируемые изопропил-1-тиогалактопиранозидом геныбактерийи. Удельная активность ксилозоизомеразы частично очищенных бесклеточных экстрактов рекомбинантов составляет соответственно 9,3 и 15,1 ед/мг 12. Наиболее близким предлагаемому по технической сущности и достигаемому результату является рекомбинантный штаммК 24 13. Для его конструирования сначала гениз 21175 в составе плазмиды 200 под контролем промотора бактериофага Т 7 клонирован в. Затем сконструирована плазмида 300 путем субклонирования генаразмером 1,2 из плазмиды 200 в плазмиду 4090, содержащую селективные маркеры, сайт инициации репликации и еЕ-промотор. Далее гибридная плазмида путем трансформации протопластов передается в 24. Рекомбинантный штамм 300 характеризуется высоким уровнем синтеза внутриклеточной ксилозоизомеразы, составляющим соответственно 6-17 и 10-24 ед/г биомассы, и продуктивностью, достигающей 1570-2440 М/минл. К недостаткам указанного штамма следует отнести длительностьпроцесса периодического выращивания продуцента, составляющего как минимум 96 ч, включая 48 ч, необходимых для приготовления посевного материала, и как минимум 48 ч собственно процесса культивирования, необходимых для биосинтеза ксилозоизомеразы 2 12214 1 2009.08.30 обязательность введения в среду для приготовления посевного материала селективного агента - антибиотика тиострептона (5 мг/мл), обеспечивающего преобладание в популяции глубинной культуры клеток рекомбинантного штамма, которые наследуют детерминирующую синтез ксилозоизомеразы гибридную плазмиду нуждаемость штамма в интенсивной (1,6 л/лмин) аэрации для реализации высокого уровня продукции фермента необходимость введения в емкости для культивирования штамма механических отбойников с целью гомогенизации и частичного разрушения растущей биомассы сложность процедуры выделения ксилозоизомеразы, включающей деструкцию клеток штамма-продуцента с использованием дорогостоящего фермента лизоцима в количестве 2 мг/мл, что требует дальнейшей очистки фермента, предназначенного для использования в пищевой промышленности, от балластного белка экстракцию фермента в буферном растворе, содержащем ЭДТА и 4, при перемешивании в течение 30 мин при 37 С ультрацентрифугирование экстракта ксилозоизомеразы при 16000 в течение 30 мин при комнатной температуре для удаления нерастворимых остатков клеточных стенок штамма-продуцента. Задачей изобретения является конструирование плазмиды, детерминирующей синтез ксилозоизомеразы в клетках содержащих их рекомбинантных бактерий, и создание нового высокоактивного штамма - продуцента ксилозоизомеразы, характеризующегося низким значением времени генерации, непродолжительным временем культивирования, стабильным наследованием инкорпорированной плазмиды в неселективных условиях, высоким уровнем экспрессии клонированного гена и, следовательно, высокой продуктивностью,умеренной зависимостью роста и синтеза фермента от аэрации питательной среды, простотой выделения фермента из клеток бактерий. Плазмида и штамм, соответствующие указанным критериям, получены в результате генно-инженерных манипуляций, включающих конструирование гибридной плазмиды 24, имеющей размер 6439 п.н. и состоящей из / фрагмента ДНК плазмиды рЕТ 24, содержащего промотор, терминатор транскрипции РНК-полимеразы бактериофага Т 7 и ген устойчивости к канамицину, а также из / фрагмента ДНК, содержащего ген ксилозоизомеразы актинобактерийБИМ В-392 Д (фиг. 1 и 2) перенос гибридной плазмиды 24 в клетки-21(3) с получением рекомбинантного штамма Е.-21(3)/24, обеспечивающего продукцию ксилозоизомеразы в количестве не менее 50 синтезированного клеткой белка (фиг. 3). Полученный штамм-21(3)/24 прошел проверку на безвредность и депонирован в Коллекции непатогенных микроорганизмов ГНУ Институт микробиологии НАН Беларуси под регистрационным номером БИМ В-427 Д. Штамм также поддерживается в лаборатории ферментов ГНУ Институт микробиологии НАН Беларуси. Штамм без потери культурально-морфологических и физиолого-биохимических свойств,в том числе и ксилозоизомеразной активности, хранится при 4-6 С нижеуказанными способами ежемесячными периодическими пересевами на агаризованную среду Луриа-Бертани, содержащую (в г/л) пептон - 10,0, дрожжевой экстракт - 5,0,- 10,0, агар-агар 15,0, в которую добавлено 20 ед/мл канамицина ежегодными периодическими пересевами на агаризованную -среду под слой минерального (вазелинового) масла лиофилизацией бактериальных клеток, выращенных на -г при 37 С в течение 1-2 сут криоконсервацией при -70 С клеток бактерий, суспендированных в 10 -ном растворе глицерина. 3 12214 1 2009.08.30 Культурально-морфологические свойства. Штамм относится к группе энтеробактерий. На -г образует полупрозрачные светло-серые колонии округлой формы диаметром 2-3 мм. Поверхность колоний гладкая, блестящая, край ровный, структура гомогенная. При росте в жидкой -среде образуют интенсивную ровную муть. Клетки палочковидной формы, грамотрицательные, неспороносные. Физиолого-биохимические признаки. Факультативный анаэроб, оптимальные условия роста - рН 7,0 и температура 37 С. Хемоорганотроф. В качестве источника азота использует минеральные соединения в аммонийной форме, а также органические вещества - пептон, триптон, дрожжевой экстракт, аминокислоты и т.д. В качестве источника углерода использует аминокислоты,глицерин, углеводы. Устойчивость к антибиотикам. Клетки проявляют устойчивость к канамицину в концентрации 20 ед/мл, обусловленную наличием локализованного на плазмиде гена устойчивости. Продукт экспрессии генаизБИМ В-391 Д в клетках Е.(1 мМ) или лактозой (0,001 ) составляет не менее 50 совокупного синтезированного белка. Реакционная смесь для определения ксилозоизомеразы содержала 0,2 мл 1 М раствора субстрата, 0,5 мл 0,2 М К,а-фосфатного буфера, рН 7,8 0,1 мл 0,1 М 472 20 мг клеток или 0,2 мл бесклеточного экстракта и дистиллированную воду до объема 2 мл. Длительность реакции изомеризации - 1 ч при 70 С. За единицу ксилозоизомеразной активности принимали такое количество фермента,которое за 1 мин трансформирует 1 мкмоль альдосахара в его кетоформу в описанных выше условиях. Продуктивность штамма выражали в единицах активности в расчете на 1 л культуральной жидкости (ед/л, или М/минл), удельную активность фермента - в единицах на 1 мг белка (ед/мг). Количественное определение альдосахаров проводили цистеин-карбазольным методом 14. Суть изобретения иллюстрируется следующими примерами. Пример 1. Конструирование плазмиды 24 и создание рекомбинантного штамма-21(3)/24 осуществляли следующим способом. Фрагменты рестрицированной ферментомхромосомной ДНКБИМ В-391 Д размером 2-6 т.п.н. выделяли и лигировали с вектором 18, обработанным тем же ферментом. Полученной смесью трансформировали компетентные клетки Е.НВ 101 (у-). Отбирали клоны, которые наследовали гибридную плазмиду 18 и после трех последовательных рассевов до изолированных колоний сохраняли способность к росту на минимальной среде, содержащей (в г/л) цитрат натрия - 0,55, ксилоза - 5,0,К 2 НР 4 - 10,5, КН 2 Р 4 - 4,5, (4)24 - 1,0, 472 - 0,5, ампициллин - 0,1. Исходный рН питательных сред - 6,8. Согласно результатам рестрикционного анализа, плазмидная ДНК отобранных клонов содержала вставочный фрагмент размером 2,5 т.п.н., а клетки рекомбинантных бактерий Е.101, наследующие плазмиды со вставкой, приобретали свойство синтезировать каталитически активный ферментный белок. Полученные результаты свидетельствуют о наличии генаизБИМ В-391 Д в составе клонированного в клетках Е.101 фрагмента ДНК. На первом этапе клонирования в векторе рЕТ 24 ген ксилозоизомеразы бактерийБИМ В-391 Д амплифицирован с использованием следующих праймеров(- )(- ). 4 12214 1 2009.08.30 Амплифицированный фрагмент для исключения вероятности клонирования неактивного гена встраивали в вектор 18 по сайтам рестрикцииис получением плазмиды рР 4. После установления методом секвенирующего анализа идентичности нуклеотидных последовательностей амплифицированного фрагмента ДНК иизБИМ В-391 Д ген ксилозоизомеразы переклонировали по указанным сайтам в вектор экспрессии рЕТ 24. Трансформацию штамма Е.-21(3) осуществляли согласно общепринятым методам. Для этого бактериями культивировали в -среде при 37 С до достижения величины оптической плотности (ОП 600) в пределах 0,3-0,4. Отделенные центрифугированием клетки бактерий ресуспендировали в охлажденном 0,1 М растворе СаС 12. Плазмидную ДНК добавляли к суспензии компетентных клеток, смесь аккуратно смешивали, разбавляли-средой, содержащей 20 мМ глюкозы, и, аэрируя, инкубировали при 37 С в течение 1 ч для экспрессии плазмидных генов антибиотикорезистентности. Смесь высевали на -агар,содержащий 20 ед/мл канамицина, и инкубировали до появления бактериальных колоний. Ксилозоизомеразную активность рекомбинантных клонов оценивали после их глубинного выращивания в -среде, содержащей в качестве индуктора изопропил-1-тиогалактопиранозид (1 мМ) и в качестве селективного агента канамицин (20 мг/мл). Посевным материалом служила 18-часовая культура бактерий, выращенных в среде с канамицином (20 мг/мл), в количестве 1 об. . Условия культивирования скорость вращения качалки - 180-200 об/мин температура 37 С, длительность культивирования - 6 ч. В описанных условиях продуктивность рекомбинантного штамма Е.21(3)/24 по ксилозоизомеразе составляет 1150 М/минл при удельной активности неочищенного фермента 2,25 ед/мг белка. Для достижения уровня синтеза ксилозоизомеразы, сравнимого с уровнем продукции фермента штаммом-прототипом, общая продолжительность стадии культивирования рекомбинантного штамма, включая время приготовления посевного материала (18 ч), составляет 24 ч, что, по меньшей мере, в 4 раза меньше, чем для этого необходимо штаммупрототипу. Пример 2. При исследовании времени генерации и стабильности наследования плазмиды 24 рекомбинантный штамм-21(3)/24 выращивали, как указано в примере 1, до достижения стационарной фазы роста. Затем культуру бактерий стократно разбавляли -средой, содержащей и не содержащей канамицин, и продолжали культивирование в вышеуказанных условиях, ежечасно определяя величину ее оптической плотности при длине волны 600 нм. Из построенного в полулогарифмических координатах графика зависимости оптической плотности культуры от времени культивирования находили время удвоения плотности культуры, определяемое как время генерации. Этот показатель у рекомбинантного штамма Е.-21(3)/24 при росте на-среде как в присутствии антибиотика, так и в его отсутствие составляет 35 мин. Определение стабильности наследования плазмиды 24 проводили на 20-й, 40-й,60-й, 80-й, 100-й и 120-й генерациях клеток рекомбинантного штамма Е.-21(3) / 24 путем их рассева на поверхность агаризованной -среды, содержащей и не содержащей канамицин в вышеуказанной концентрации. Стабильно высокий уровень наследования исследуемой плазмиды 24 в клетках рекомбинантного штамма выявляется, по меньшей мере, в 120-й генерации клеток, независимо от присутствия в среде селективного агента - канамицина. Косвенным подтверждением отмеченного факта является высокая продуктивность различных генераций клеток по ксилозоизомеразе 1100-1500 М/минч. Клетки штамма-прототипа, в отличие от предлагаемого нами рекомбинантного штамма, поддерживают плазмиду в течение 17 генераций, после чего наступает их лизис. 5 12214 1 2009.08.30 Пример 3. Глубинное выращивание рекомбинантного штамма-21(3) / 24 проводят, как описано в примере 1, за исключением того, что вместо изопропил-1-тиогалактопиранозида (1 мМ) в качестве индуктора генаиспользуют менее дефицитный и дорогой аналог - лактозу в количестве 0,001 , а из среды для приготовления посевного материала исключают селективный агент - канамицин. Продуктивность рекомбинантных бактерий по ксилозоизомеразе составила 1500 М/минл,удельная активность фермента - 2,0 ед/мг белка. Пример 4. Масштабирование процесса культивирования рекомбинантного штамма Е.21(3)/24 осуществляют в 30 л ферментере при соблюдении следующих условий коэффициент заполнения - 0,5 скорость подачи воздуха - 1 л/л среды в минуту скорость вращения мешалки - 200 об/мин температура культивирования - 37 С. Составы питательных сред для получения инокулюма и выращивания штамма, а также условия индукции генаи длительность культивирования аналогичны указанным в примере 3. Через 1 ч после добавления лактозы в среду культивирования у рекомбинантного штамма инициируется синтез ферментного белка, а его максимальная продуктивность достигает 2540 М/минл через три часа, что соответствует ранней стационарной фазе роста (фиг. 4). Через 7,5-8 ч от начала культивирования наступает практически полный лизис культуры. Следовательно, выделение фермента из клеток предлагаемого рекомбинантного штамма-21(3)/24, в отличие от штамма-прототипа, осуществляется спонтанно и не требует дополнительной ферментативной деструкции клеток. Пример 5. Проводят культивирование рекомбинантного штамма, как описано в примере 4, за исключением того, что интенсивность аэрации питательной среды увеличивают до 1,6 л/л среды, что соответствует оптимальным условиям для образования ксилозоизомеразы штаммом-прототипом. Продуктивность рекомбинантного штамма-21(3)/24 ксилозоизомеразе в условиях повышенной в 1,6 раза аэрации составила 2520 М/минл,что равнозначно этому показателю у культуры, выращенной в условиях ее невысокого снабжения кислородом. Пониженный уровень аэрации среды, необходимый для достижения предлагаемым штаммом-продуцентом такого же уровня продуктивности, как у штамма-прототипа, обеспечивает первому экономические преимущества. Пример 6. Проводят культивирование рекомбинантного штамма, как описано в примере 4, и получают 18 л культуральной жидкости, содержащей лизированные клетки Е. со-21(3)/24, с суммарной ксилозоизомеразной активностью 45540 ед. Для получения бесклеточного препарата ксилозоизомеразы культуральную жидкость нагревают при 70 С в течение 10 мин для удаления термолабильных белков, центрифугируют (8000 , 10 мин) и 20-кратно концентрируют через мембранные фильтры УПМ-20(Владипор, Россия). Получают бесклеточный концентрат ксилозоизомеразы в количестве 0,89 л с выходом по ферментативной активности 95 . Таким образом, заявляемый рекомбинантный штамм-21(3) / 24 БИМ В-427 Д, в течение, по меньшей мере, 120 генераций стабильно наследующий заявляемую плазмиду, по важнейшим показателям (ферментативной активности,продуктивности, технологическим свойствам) превосходит известные аналоги, что позволяет рекомендовать его для получения микробной ксилозоизомеразы, востребованной в производстве глюкозо-фруктозных сиропов и кристаллической фруктозы. 6 Индукция изопропил-1-тиогалактопиранозидом (ИПТГ) и лактозой синтеза ксилозоизомеразы рекомбинантным штаммом Е.-21(3)/24. 1 - маркер молекулярной массы 2 - белок клеток до внесения индуктора,синтез фермента клетками через один, два и четыре ч а с а и н д ук ц и и ИП Т Г (3 , 4 , 5 , с оо т в е т с т в ен н о ) лактозой (6,7, 8, соответственно) Рост рекомбинантного штамма Е.21(3)/24 и синтез ксилозоизомеразы в ферментере. 1 - биомасса, ОП 600,2 - ксилозоизомераза, ед./мл. Стрелка указывает на время внесения лактозы в среду культивирования Национальный центр интеллектуальной собственности. 220034, г. Минск, ул. Козлова, 20. 8
МПК / Метки
МПК: C12N 15/61
Метки: клетках, обеспечивающая, штамм, бим, в-427, рекомбинантный, продуцент, pet24bxyla, плазмида, штамма, escherichia, рекомбинантного, ксилозоизомеразы, синтез
Код ссылки
<a href="https://by.patents.su/8-12214-plazmida-pet24bxyla-obespechivayushhaya-sintez-ksilozoizomerazy-v-kletkah-rekombinantnogo-shtamma-escherichia-coli-bim-v-427-d-i-rekombinantnyjj-shtamm-escherichia-coli-bim-v-427-d.html" rel="bookmark" title="База патентов Беларуси">Плазмида pET24bxylA, обеспечивающая синтез ксилозоизомеразы в клетках рекомбинантного штамма Escherichia coli БИМ В-427 Д, и рекомбинантный штамм Escherichia coli БИМ В-427 Д – продуцент ксилозоизомеразы</a>
Рекомбинантная плазмидная ДНК, кодирующая препроинсулин человека и штамм Escherichia coli XLI-Blue/pInsR-продуцент препроинсулина

Номер патента: 3617
Опубликовано: 30.12.2000
Авторы: Уваров Валентин Юрьевич, Сивов Игорь Геннадьевич, Маркин Сергей Сергеевич, Семенов Михаил Петрович, Нечаев Виктор Николаевич
МПК: C12N 15/17
Метки: препроинсулина, днк, escherichia, препроинсулин, плазмидная, кодирующая, рекомбинантная, штамм, человека
Текст:
...связан через аргинин с естественной лидерной последовательностью и имеющую нуклеотидную последовательность, приведенную на фиг. 1. Фиг. 1. Участок нуклеотидной последовательности плазмиды , соответствующей гену рекомбинантного белка препроинсулина человека. Место проведения точечной замены подчеркнуто жирным шрифтом. Поставленная задача решается также созданием штамма . 1-/ - продуцента препроинсулина, характеризующегося наличием...
Штамм Escherichia coli КМИЭВ-22 – штамм-антиген

Номер патента: 10884
Опубликовано: 30.08.2008
Авторы: Андросик Николай Николаевич, Полоз Светлана Васильевна, Карпович Валентина Константиновна, Ломако Юрий Васильевич
МПК: C21N 1/20
Метки: штамм-антиген, штамм, кмиэв-22, escherichia
Текст:
...ветеринарии им. С.Н. Вышелесского Национальной академии наук Беларуси. Таксонометрическая принадлежность. Согласно определителю бактерий Берджиотносится к 5-ой группе факультативно анаэробных грамотрицательных палочек, подгруппе 1 - семейство , роду , виду. Культурально-морфологические признаки. ШтаммО 33 (КМИЭВ-22) при окраске по Граму имеет вид толстых, коротких (0,2-0,72-4 мкм) грамотрицательных, не образующих спор, палочек....
Штамм бактерий Escherichia coli, продуцирующий нуклеозидфосфотрансферазу Erwinia herbicola

Номер патента: 11916
Опубликовано: 30.06.2009
Авторы: Квач Сергей Вячеславович, ЕРОШЕВСКАЯ Людмила Анатольевна, Зинченко Анатолий Иванович
Метки: штамм, herbicola, erwinia, нуклеозидфосфотрансферазу, escherichia, бактерий, coli, продуцирующий
Текст:
...клеток. Содержание НФТ в клетках штаммаНФТ 15 составляет до 10 от суммарного содержания белка и фермент после разрушения клеток, например, ультразвуком может быть использован без дополнительной очистки или очищен до гомогенного состояния стандартными методами. Для культивированияНФТ 15 с целью получения НФТ применяют питательную среду(рН 7,0), приготовленную на водопроводной воде, следующего составатриптон ( США) - 1,0 дрожжевой экстракт (...
Штамм Escherichia coli КМИЭВ-18 – штамм-антиген

Номер патента: 10979
Опубликовано: 30.08.2008
Авторы: Ломако Юрий Васильевич, Полоз Светлана Васильевна, Андросик Николай Николаевич, Карпович Валентина Константиновна
МПК: C12N 1/20
Метки: штамм-антиген, кмиэв-18, штамм, escherichia
Текст:
...выделен штамм 18- (КМИЭВ-18), отличающий наличием в антигенной структуре соматического антигена 018, который был определен в реакции агглютинации с монорецепторной сывороткой. Штамм бактерий 18-М депонирован в коллекции РНИУП Институт экспериментальной ветеринарии им. С.Н.Вышелесского Национальной академии наук Беларуси под номером КМИЭВ-18. Таксонометрическая принадлежность. Согласно определителю бактерий Берджиотносится к 5-й группе...
Штамм Escherichia coli КМИЭВ-38 – штамм-антиген

Номер патента: 10978
Опубликовано: 30.08.2008
Авторы: Андросик Николай Николаевич, Полоз Светлана Васильевна, Карпович Валентина Константиновна, Ломако Юрий Васильевич
МПК: C12N 1/20
Метки: escherichia, кмиэв-38, штамм, штамм-антиген
Текст:
...имеет вид тонких, коротких (0,2-0,72-4 мкм) грамотрицательных палочек. Физиолого-биохимические признаки. Факультативный анаэроб. Растет в МПБ, в бульоне на основе перевара Хоттингера с интенсивным помутнением среды и образованием на дне пробирки осадка, легко разбивающегося при встряхивании. Культура на плотных питательных средах формирует круглые, блестящие, выпуклые колонии с приподнятым,более оптически плотным центром. На среде Эндо...
Предыдущий патент: Индуктор созревания моноцитарных дендритных клеток in vitro
Следующий патент: Мельница пружинная
Случайный патент: Градирня вентиляторная