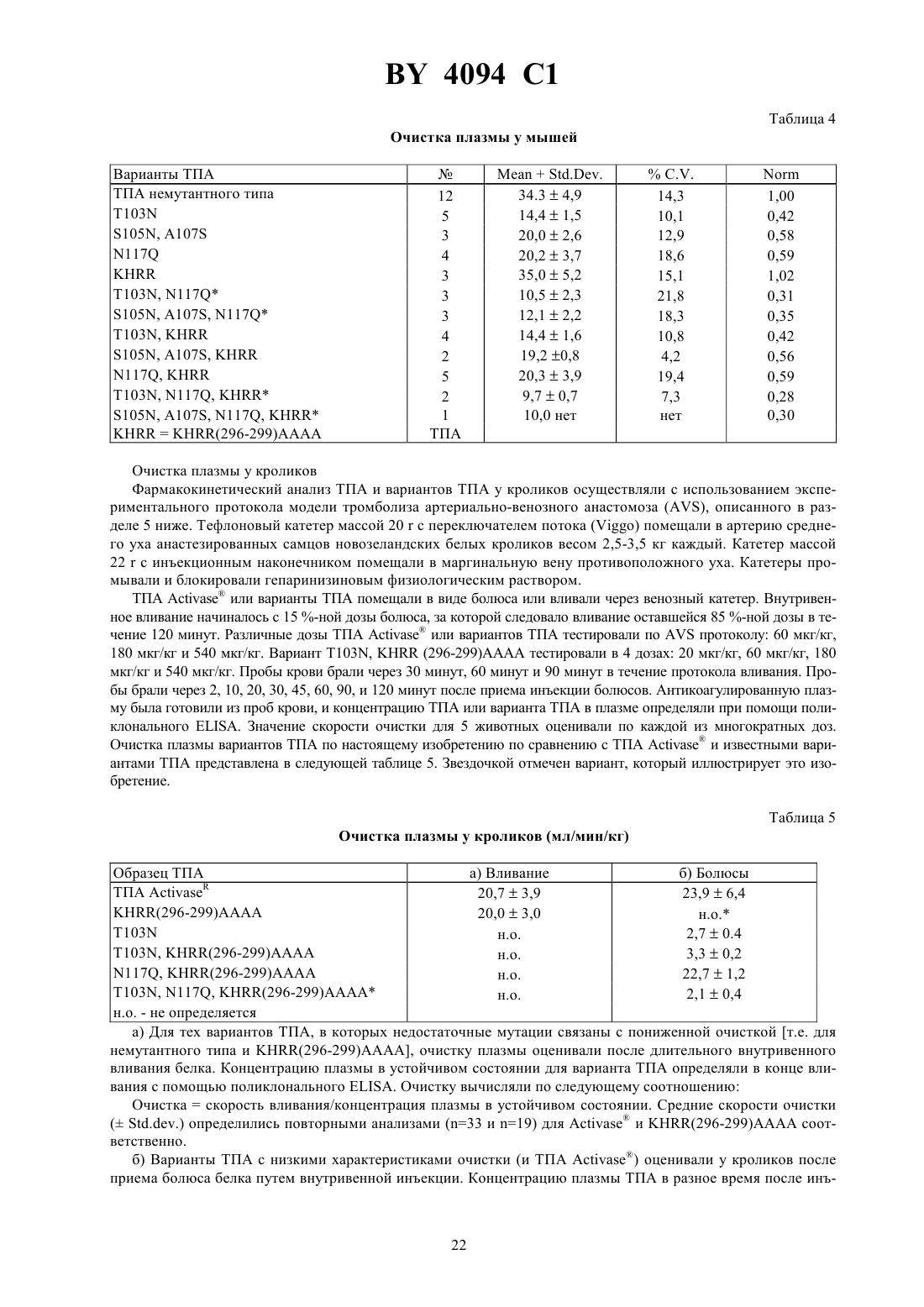
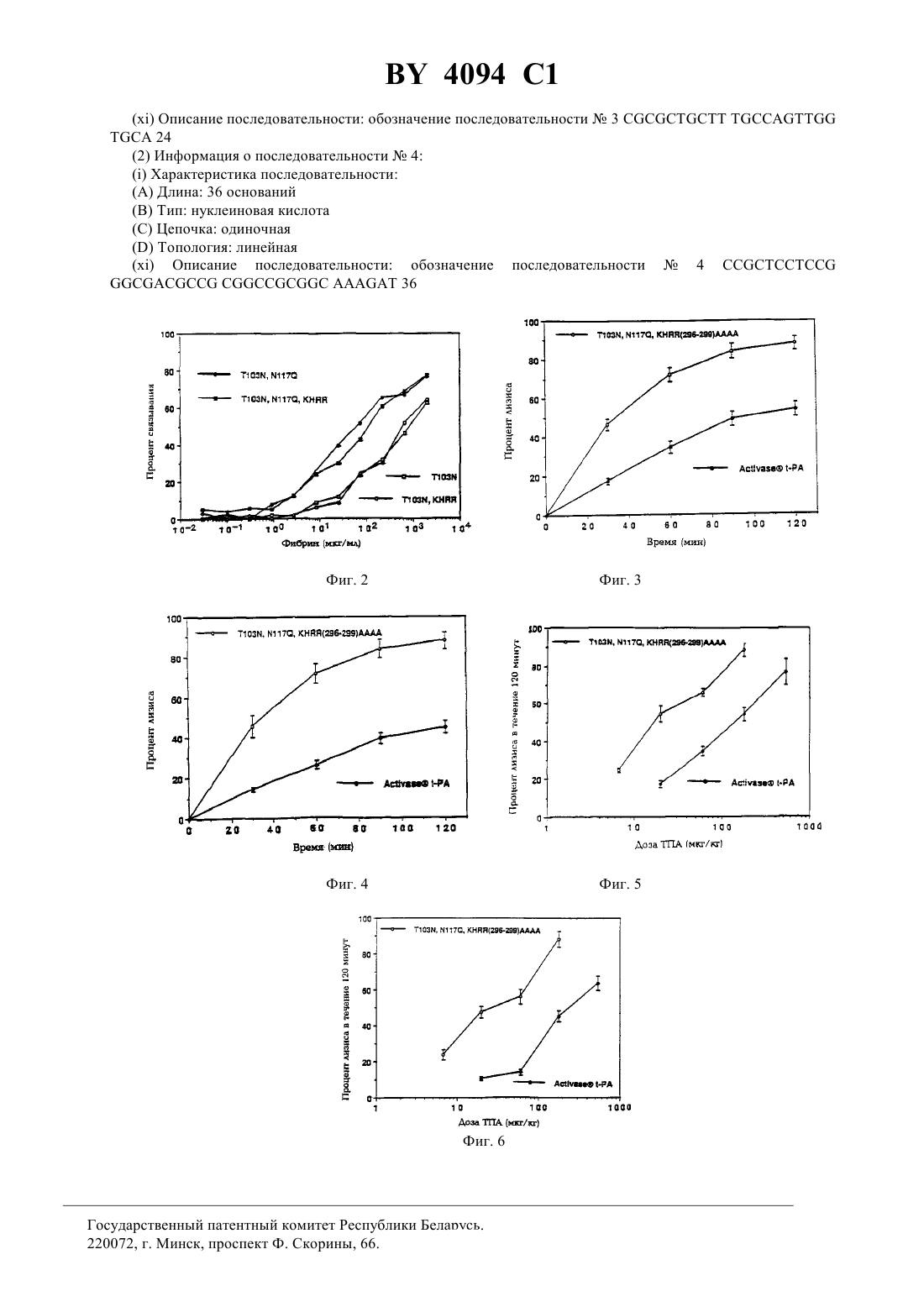
Варианты человеческого тканевого плазминогенного активатора (ТПА), последовательности ДНК, кодирующие варианты ТПА, составы, способы лечения и способ восстановления свойства связывания фибрина с ТПА
Номер патента: 4094
Опубликовано: 30.09.2001
Авторы: ПАОНИ, Николас, Ф., БЕННЕТТ, Уиллиам, Ф., КЕЙТ, Брюс, Алан
























Текст
ГОСУДАРСТВЕННЫЙ ПАТЕНТНЫЙ КОМИТЕТ РЕСПУБЛИКИ БЕЛАРУСЬ ВАРИАНТЫ ЧЕЛОВЕЧЕСКОГО ТКАНЕВОГО ПЛАЗМИНОГЕННОГО АКТИВАТОРА (ТПА), ПОСЛЕДОВАТЕЛЬНОСТИ ДНК, КОДИРУЮЩИЕ ВАРИАНТЫ ТПА, СОСТАВЫ, СПОСОБЫ ЛЕЧЕНИЯ И СПОСОБ ВОССТАНОВЛЕНИЯ СВОЙСТВА СВЯЗЫВАНИЯ ФИБРИНА С ТПА(57) 1. Вариант человеческого тканевого плазминогенного активатора (ТПА), отличающийся тем, что он гликозилирован в любой из позиций 103-105 и лишен функциональной углеводной структуры в позиции 117 последовательности аминокислот человеческого ТПА немутантного типа, причем указанный вариант проявляет расширенный циркулирующий период полураспада и в значительной степени сохраненное связывание фибрина или повышенную фибринолитическую активностьпо сравнению с человеческим ТПА немутантного типа. 2. Вариант ТПА по п. 1, отличающийся тем, что он имеет расширенный циркулирующий период полураспада и в значительной степени сохраненное связывание фибрина по сравнению с человеческим ТПА немутантного типа. 3. Вариант ТПА по п. 2, отличающийся тем, что он имеет повышенное связывание фибрина по сравнению с вариантом человеческого ТПА, гликозилированным в любой из позиций 103-105 и имеющим функциональную углеводную структуру в позиции 117 человеческого ТПА немутантного типа. 4. Вариант ТПА по п. 1, отличающийся тем, что он имеет повышенную фибринолитическую активностьпо сравнению с человеческим ТПА немутантного типа. Фиг. 1 5. Вариант ТПА по п. 1, отличающийся тем, что он гликозилирован в позиции 103 последовательности аминокислот человеческого ТПА немутантного типа. 4094 1 6. Вариант ТПА по п. 1, отличающийся тем, что он гликозилирован в позиции 105 последовательности аминокислот человеческого ТПА немутантного типа. 7. Вариант ТПА по п. 1, отличающийся тем, что он гликозилирован в позициях 103-105 через -связь. 8. Вариант ТПА по п. 1, отличающийся тем, что он гликозилирован в позициях 103-105 через 0-связь. 9. Вариант ТПА по п. 7, отличающийся тем, что он содержит аспарагин, как часть или -трипептидной последовательности, где Х - любая аминокислота, за исключением пролина, в позиции 103 последовательности аминокислот человеческого ТПА немутантного типа. 10. Вариант ТПА по п. 9, отличающийся тем, что он содержит аспарагин в позиции 103, триптофан в позиции 104 и серин в позиции 105 аминокислотной последовательности человеческого ТПА немутантного типа. 11. Вариант ТПА по п. 7, отличающийся тем, что он содержит аспарагин как часть или -трипептидной последовательности, где Х - любая аминокислота, за исключением пролина, в позиции 105 последовательности аминокислот человеческого ТПА немутантного типа. 12. Вариант ТПА по п. 11, отличающийся тем, что он содержит аспарагин в позиции 105, треонин в позиции 106 и серин в позиции 107 последовательности аминокислот человеческого ТПА немутантного типа. 13. Вариант ТПА по п. 1, отличающийся тем, что он содержит другую аминокислоту, чем аспарагин, в позиции 117 последовательности аминокислот человеческого ТПА немутантного типа. 14. Вариант ТПА по п. 13, отличающийся тем, что он содержит глутамин в позиции 117 последовательности аминокислот человеческого ТПА немутантного типа. 15. Вариант ТПА по п. 13, отличающийся тем, что он дополнительно содержит аспарагин как часть - или трипептидной последовательности, где Х - любая аминокислота, за исключением пролина, в позиции 103 последовательности аминокислот человеческого ТПА немутантного типа. 16. Вариант ТПА по п. 13, отличающийся тем, что он дополнительно содержит аспарагин как часть - или трипептидной последовательности, где Х - любая аминокислота, за исключением пролина, в позиции 105 последовательности аминокислот человеческого ТПА немутантного типа. 17. Вариант ТПА по п. 16, отличающийся тем, что он дополнительно содержит серин в позиции 107 последовательности аминокислот человеческого ТПА немутантного типа. 18. Вариант ТПА по п. 1, отличающийся тем, что он имеет повышенную фибриноспецифичность по сравнению с человеческим ТПА немутантного типа. 19. Вариант ТПА по п. 18, отличающийся тем, что указанная фибриноспецифичность достигается путем иного изменения, чем замещение аспарагиновой кислоты аргинином в позиции 299, в пределах аминокислотной области 296-302 или 274-277 последовательности аминокислот человеческого ТПА немутантного типа. 20. Вариант ТПА по п. 19, отличающийся тем, что указанное изменение находится в области 296-300 последовательности аминокислот человеческого ТПА немутантного типа. 21. Вариант ТПА по п. 20, отличающийся тем, что указанное изменение находится в одной или более позициях 296, 297, 298 и 299 последовательности аминокислот человеческого ТПА немутантного типа. 22. Вариант ТПА по п. 21, отличающийся тем, что указанное изменение является замещением аланина каждой из аминокислот лизином, гистидином, аргинином в позициях 296, 297, 298 и 299 последовательности аминокислот человеческого ТПА немутантного типа. 23. Вариант ТПА по п. 19, отличающийся тем, что он содержит аспарагин как часть или - трипептидной последовательности в позиции 103 и аминокислоту иную, чем аспарагин, в позиции 117 последовательности аминокислот человеческого ТПА немутантного типа. 24. Вариант ТПА по п. 23, отличающийся тем, что он содержит аспарагин в позиции 103, триптофан в позиции 104, серин в позиции 105 и глутамин в позиции 117 последовательности аминокислот человеческого ТПА немутантного типа. 25. Вариант ТПА по п. 19, отличающийся тем, что он содержит аспарагин как часть или - трипептидной последовательности в позиции 105 и аминокислоту иную, чем аспарагин, в позиции 117 последовательности аминокислот человеческого ТПА немутантного типа. 26. Вариант ТПА по п. 25, отличающийся тем, что он содержит аспарагин в позиции 105, треонин в позиции 106, серин в позиции 107 и глутамин в позиции 117 последовательности аминокислот человеческого ТПА немутантного типа. 27. Вариант ТПА по п. 1, отличающийся тем, что он выбран из группы, состоящей из вариантов человеческого ТПА, содержащих следующие мутации 103, 117 105, 107, 117 103, 117,К 296 А, Н 297 А, 298, 299 105, 107, 117, К 296 А, Н 297 А, 298, 299 103, 117,298, 299 105, 107, 117, 298, 299 103, 117, 296, 297, 301 105, 107,117, 296, 297, 301 103, 117, 274, 275, 276, К 277 Т 105, 107, 117, 274,275, 276, К 277 Т, гдеобозначает любую аминокислоту, за исключением аспарагина . 28. Последовательность ДНК, кодирующая вариант ТПА по п. 1, содержащая нуклеотиды. 2 4094 1 29. Последовательность ДНК, кодирующая вариант ТПА по п. 1, содержащая нуклеотиды. 30. Последовательность ДНК, кодирующая вариант ТПА по п. , содержащая нуклеотиды. 31. Последовательность ДНК, кодирующая вариант ТПА по п. 1, содержащая нуклеотиды. 32. Состав для лечения сосудистого нарушения или заболевания, содержащий терапевтически активное вещество и фармацевтически приемлемый носитель, отличающийся тем, что в качестве терапевтически активного вещества он содержит вариант ТПА по п. 1 в эффективном количестве. 33. Способ лечения сосудистого нарушения или заболевания у млекопитающих, отличающийся тем, что млекопитающему вводят эффективное количество состава по п. 32. 34. Состав для предотвращения отложения фибрина или образования или преобразования адгезии, содержащий терапевтически активное вещество и фармацевтически приемлемый носитель, отличающийся тем, что в качестве терапевтически активного вещества он содержит вариант ТПА по п. 1 в эффективном количестве. 35. Способ лечения млекопитающего для предотвращения отложения фибрина или образования или преобразования адгезии, отличающийся тем, что в сайт потенциального образования фибрина или адгезии у млекопитающего вводят эффективное количество состава по п. 34. 37. Способ восстановления свойства связывания фибрина с человеческим ТПА немутантного типа, отличающийся тем, что ТПА редуцируют в результате гликозилирования в любой из аминокислотных позиций 103-105 и удаляют функциональную углеводную структуру в аминокислотной позиции 117.(56)89/01947,1989.91/05418,1991.89/03755,1989.89/00705,1989. Настоящее изобретение относится к недифференцированным клеткам человека, а именно к тканевому плазминогенному активатору (ТПА), и к ДНК, кодирующей ТПА, а также к составам и способам лечения, и к способам восстановления свойства связывания фибрина с ТПА. Тканевый плазминогенный активатор (ТПА) является полидоменной сериновой протеазой, физиологическая роль которой состоит в преобразовании плазминогена в плазмин, а, следовательно, в возбуждении или ускорении процесса фибринолиза. Вначале клинический интерес к ТПА возник благодаря его относительно высокой активности в присутствии фибрина по сравнению с его отсутствием. ТПА немутантного типа является слабым ферментом в отсутствии фибрина, но присутствие фибрина резко увеличивает его способность к активизации плазминогена. Без стимуляции каталитическая эффективность (постояннаястепени катализа/постоянная Михаэлиса(К меланомы или рекомбинантного человеческого ТПА (ТПА ) для активизации плазминогена равна приблизительно 1 нМ-1 с-1 в то время, как в присутствии фибрина или продуктов распада фибрина, эта эффективность (постоянная псевдостепени) повышается на несколько сотен порядков. Считают, что это необычное биохимическое свойство ТПА переходит клинически в тромболитический продукт, который менее пригоден, чем нефибриноспецифические продукты, растворяющие тромбы, (такие, как стрептокиназа или урокиназа) для стимуляции систематической плазминогенной активации , В.Е..,69,983-990 (1984). Рекомбинантный человеческий ТПА используется терапевтически как фибринолитический агент при лечении острого инфаркта миокарда и легочной эмболии, причем оба состояния обычно возникают от закупорки кровеносного сосуда фибриносодержащим тромбом. Кроме этой неожиданной фибриноспецифичности, ТПА проявляет еще некоторые отличительные свойства а) ТПА отличается от большинства сериновых протеаз тем, что молекула, имеющая форму одной цепочки, проявляет заметную ферментативную активность. По отношению к некоторым небольшим субстратам и по отношению к плазминогену в отсутствии фибрина двухцепочный ТПА имеет большую активность, чем одноцепочный. Однако, в присутствии фибрина обе формы ТПА одинаково активныа 1 257, 2920-5 (1982)., . . 64, 61-8 (1990)., . . . 266, 5191-5201 (1991). Большинство других сериновых протеаз существует как зимогены и для реализации полной ферментативной активности нуждаются в протеолитическом расщеплении до двухцепочной формы. б) Действие ТПАиможет подавляться серпином, РА-1 , ., ,84, 586-591 (1989) , .., . . . 259, 3644 3647 (1984). в) В -диапазоне ТПА связанс фибрином с помощью . 4094 1 д) ТПА быстро очищается. Медиатором данного процесса являются один или более рецепторов в печени , .., . . 39, 511-521 (1985) , .., . . 53, 287-303 (1989), ., . . . 264,7228-7235 (1989). В значительной степени чистая форма ТПА была впервые получена из натурального источника и испытана на активностьа 1., патент США 4 752 603, выданный 21 июня 1988 г. (см. также.,, 256 7035 1981).., (, 301214 1983) определили последовательность ДНК в ТПА и установили последовательность аминокислот этой последовательности ДНК (см. патент США 4766075, выданный 23 августа 1988 г). Человеческий ТПА немутантного типа имеет потенциальные -связанные гликозилированные сайты в позициях аминокислот 117, 184, 218 и 448. Известно, что рекомбинантный человеческий ТПА (ТПА), полученный в результате экспрессии в клетки СНО (яичника китайского хомячка), содержит приблизительно 7 вес.углевода, состоящего из олигосахарида с высоким содержанием маннозы в позиции 117 и сложных олигосахаридов в позициях -184 и -448 , .,1986 551-562. Позиция 218, как было обнаружено, не была гликозилирована в природном ТПА. Оказалось, что сайты 117 и 448, всегда гликозилированы, в то время как в сайте 184 гликозилируется около 50 молекул. Молекулы ТПА, которые гликозилированы в позиции 184, называются типомТПА, а молекулы, которые не гликозилированы в позиции 184, называются типомТПА. Самый исчерпывающий анализ углеводных структур, полученных из клеток СНО человеческого ТПА был проведен., . . ., 264 (24) 14100-14111 (1989), который показал, что в белке можно обнаружить по крайней мере 17 различных углеводных структур, связанных с(аспирином). Последние имеют диапазон от структур с высоком содержанием маннозы в позиции 117 до ди-, три- и тетра- антенных структур типа-ацетиллактозамина в позициях 184 и 448. Обнаружено, что типи типТПА были -гликозилированы идентичным образом в позициях -117 и -448, выделенных от линии той же клетки. Более подробно см. также ,В..,28, 7644-7662 (1989). Анализ последовательности ТПА идентифицировал как молекулу, состоящую из 5 доменов. В каждом домене были определены гомологические структурные или функциональные зоны по отношению к другим протеинам, таким как трипсин, химотрипсин, плазминоген, протромбин, фибронектин и фактор роста эпидермиса . Были указаны номера терминальных участков последовательности аминокислот ТПА, в которых начинались эти домены, такдомен- примерно от аминокислоты 1 до аминокислоты 44, домен фактора роста- примерно от аминокислоты 45 до аминокислоты 91 (на основе гомологии с ) домен -1 (К 1) - примерно от аминокислоты 92 до аминокислоты 173, домен -2 (К 2) - примерно от аминокислоты 180 до аминокислоты 261, а домен сериновой протеазы (Р) - примерно от аминокислоты 264 до карбоксильной терминали в аминокислоте 527. Эти домены расположены в основном смежно друг с другом и связаны короткими линкерными участками. Эти линкерные участки приводят к общему количеству аминокислот сформированного полипептида, равному 527, хотя три дополнительных остатка (-) редко находятся в терминальном участке аминокислот. Обычно считают, что этот дополнительный трипептид образуется в результате незавершенного предшествующего процессинга, и неизвестно, кодирует ли он функционирование. Природный ТПА может быть расщеплен между позицией 275 и позицией 276 (расположенными в домене сериновой протеазы) с образованием двухцепочной формы молекулы. Каждый домен по-разному участвует в общих биологически существенных свойствах молекулы ТПА. Исследования по делеции доменов показывают, что потеря доменов , фактора роста или -2 вызывает более низкое специфическое связывание варианта ТПА с фибрином, ., .. . .83, 4760-4677 (1986) , .,.5, 3525-30 (1986), однако, более поздние результаты, полученные с замещенными мутантами, показывают, что домен -2 меньше участвует в связывании фибрина, чем ожидалось ранее ., . 266 5191-5201 (1991). Исследования по делеции доменов обнаружили участие в очистке печени доменови фактора роста.,71, 216-219 (1988)., . . . 263, 3971-3978 (1988)., . . 50, 33-41 (1988).,2, 30 (1988).,73 1842-1850 (1989)., 263, 1599-1602 (1988). Домен -2 отвечает за связывание с лизином. Было показано, что домен сериновой протеазы отвечает за ферментативную активность ТПА и содержит специфические зоны, в которых мутации влияют как на связывание с фибрином, так и на фибриноспецифичность (возможность прямых взаимодействий с фибрином), и другие зоны, в которых изменяется только фибриноспецифичность(возможность косвенных взаимодействий с фибрином)., 1991, выше. Исследования с мутантами, полученными в результате изменений, направленных на сайт, показывают участие гликозилирования ТПА в очистке., / 5, 953-958 (1987)., / 6. 734 (1988). Относительно быстрая очистка от плазмы человеческого ТПА немутантного типа является полезным для пациентов, которым крайне необходимо вмешательство после тромболиза и требуется длительное внутривенное введение для сохранения терапевтических уровней ТПА в крови. Рекомендуемая общая доза ТПАдля тромболитической терапии при острой легочной эмболии для взрослых пациентов составляет 4 4094 1 100 мг при длительном внутривенном вливании в течение 2 часов. Производные ТПА с более длинным периодом полураспада плазмы (более медленной очисткой) могли бы вводить в виде инъекций болюса и иметь при этом более высокую концентрацию плазмы, чем это могло быть получено длительным введением ТПА немутантного типа, что может вызвать снижение эффективной дозы. Мутанты ТПА с более медленной очисткой могли бы обеспечить конкретные преимущества при лечении таких состояний, как тромбоз глубоких кровеносных сосудов, лечении, сопровождающем повторную перфузию жертвы инфаркта, лечении легочной эмболии или лечении периферического артериального тромбоза (заболевания периферической сосудистой системы). Необходимость в вариантах ТПА, которые могут вводить в форме болюса, объясняется в последнее время интересом к введению болюса человеческого ТПА немутантного типа (особенно в форме одной цепочки) с целью обеспечения раннего раскрытия коронарных артерий, связанных с инфарктом, и содействия спасения от миокардита.,14, 1566-1569 (1989) ,14, 1566-1569 (1989)., 65, 11051-1056 (1990) , ., 68, 1570-1574 (1991). Результаты недавних клинических исследований показывают, что внутривенное введение в виде болюса человеческого ТПА немутантного типа не только ускоряет начало тромболитической терапии, но и быстрое достижение относительно высокой концентрации ТПА также усиливает тромболиз , ., выше, , , .. . . 15 922-924 (1990). Варианты ТПА с пониженной очисткой были получены путем делеции отдельных аминокислот, частичных доменов или полных доменов из молекулы. В следующих публикациях представлены попытки уменьшить скорость очистки ТПА немутантного типа путем делеции части или всего домена фактора роста и/или домена , по выбору скомбинированных с другими мутациями. ., 1988, выше.,. 63, 54-59 (1990), 1988, выше., 1988, выше.,81, 1362-73 (1990)., ., 11 468 (1988) .,3091 (1990).,4, 131-140 (1990) патент США 4 935 237 (выдан 19 июня 1990 г.) Европейская патентная заявка 241 208 (опубликована 14 октября 1987 г.) Европейская патентная заявка 240 334 (опубликована 7 октября 1987 г.). Вариант ТПА с удвоенным доменом -2 с пониженной очисткой плазмы был раскрыт в., . . 65, 174-180 (1991). Множество вариантов ТПА с замещением аминокислот оценивали по их способности к уменьшению скорости очистки и/или увеличению периода полураспада ТПА. Стало известно, что замещение в области аминокислот 63-72 (в частности, в позициях 67 и 68) и 42-49, увеличивает период полураспада плазмы человеческого ТПА немутантного типа (см.89/12681, опубликованную 28 декабря 1989 г., и., 265, 5540 (1990). Было обнаружено, что природный сформированный ТПА, у которого в позиции 275 аргинин замещен глутаминовой кислотой, имеет скорость очистки почти в 2 раза меньше, чем человеческий ТПА немутантного типа., . ., 58 491 (1987). Другой подход к уменьшению скорости очистки и/или расширению периода полураспада ТПА состоял в создании комплекса молекулы ТПА со второй молекулой. Из Европейской патентной заявки 304 311(опубликована 22 февраля 1989 г.) стало известно, что конъюгат ТПА и полиэтиленгликоля уменьшает скорость очистки ТПА. Обнаружили, что моноклональное антитело к ТПА увеличивает функциональный период полураспада ТПАбез снижения его активности (см. Европейскую патентную заявку 339 505,опубликованную 2 ноября 1989 г.). Также исследовали вовлечение углеводов в очистку ТПА. Были созданы и испытаны варианты ТПА с различными показателями углеводов в отличие от таких же вариантов человеческого ТПА немутантного типа. В качестве примеров такого подхода одна или более позиций 60, 64, 65, 66, 67, 78, 79, 80, 81, 82, 103, 105,107 и 250 были замещены соответствующими аминокислотами для получения молекул с сайтами гликозилирования в этих остатках или вблизи некоторых из них (см.89/11531, опубликованную 30 ноября 1989). Из этих вариантов ТПА в большой степени гликозилированный мутант ТПА 103, например, имеет скорость очистки на 5 порядков ниже, чем природный ТПА. В другой публикации внимание сконцентрировано на преобразовании сайтов гликозилирования ТПА немутантного типа в негликозилированные сайты. Обнаружили, что негликозилированный вариант ТПА, содержащий домен -2 и протеазный домены имеет более медленную скорость очистки плазмы, чем ТПА немутантного типа.,4, (. 3)9 ( 26) (1990.. .., 60 255 (1988) селективно удаляли олигосахаридные остатки из молекул ТПА и показали, что удаление этих остатков уменьшает скорость очистки ТПА. Эти исследователи и. (1987), выше (1988),выше также получили вариант ТПА - 117 (в котором аспарагин в позиции 117 человеческого ТПА немутантного типа был замещен глутамином) для предотвращения гликозилирования в позиции 117. Этот вариант, подобно варианту, полученному путем ферментативного удаления олигосахарида с высоком содержанием маннозы в этой позиции, проявляет скорость очистки почти на два порядка ниже, чем человеческий ТПА немутантного типа. См. также Европейскую патентную заявку 238 304, опубликованную 23 сентября 1987 г., и Европейскую патентную заявку 227 462, опубликованную 1 июля 1987 г. 5 4094 1 Следующие варианты гликозилирования человеческого ТПА имеющие пониженные скорости очистки,были описаны в следующих публикациях., выше (42, Н 44 Е, 117 -)., (1988) выше(6-186) 11 - и(6-186) 117, 184, 448 -., 2,611 (1989) (117, 184 -). Мы обнаружили, что расширение циркулирующего периода полураспада человеческого ТПА немутантного типа путем увеличения дополнительного гликозилирования в позициях аминокислот 103-105 также сопровождалось потерями специфичности связывания с фибрином и/или активности лизиса тромбов в плазме. Например, замена треонина на аспарагин в позиции аминокислот 103 человеческого ТПА немутантного типа снижает скорость очистки почти на 5 порядков, а также приводит к потере функции ТПА, выражающейся в том, что специфичность связывания с фибрином и активность ТПА значительно понижаются. Хотя благодаря этой пониженной скорости очистки концентрация плазмы при этом варианте гликозилирования была на 4-5 порядков выше, чем для эквивалентной дозы ТПА , в результате его пониженной активности улучшение эффективности или фибринолитической активностибыло мало. Задачей изобретения является получение вариантов ТПА с улучшенными фармакологическими свойствами с восстановленной способностью связывания с фибрином, а также получение составов и способов лечения заболеваний у млекопитающих. В результате изобретения получен вариант человеческого тканевого плазминогенного активатора (ТПА),гликозилированный в любой из позиций 103-105 и лишенный функциональной углеводной структуры в позиции 117 последовательности аминокислот человеческого ТПА немутантного типа, причем указанный вариант проявляет расширенный циркулирующий период полураспада и в значительной степени сохраненное связывание фибрина или повышенную фибринолитическую активностьпо сравнению с человеческим ТПА немутантного типа. Один из вариантов ТПА имеет расширенный циркулирующий период полураспада и в значительной степени сохраненное связывание фибрина по сравнению с человеческим ТПА немутантного типа. Предпочтительно вариант ТПА имеет повышенное связывание фибрина по сравнению с вариантом человеческого ТПА, гликозилированным в любой из позиций 103-105 и имеющим функциональную углеводную структуру в позиции 117 человеческого ТПА немутантного типа. Один из вариантов ТПА имеет повышенную фибринолитическую активностьпо сравнению с человеческим ТПА немутантного типа. Один вариант ТПА гликозилирован в позиции 103 последовательности аминокислот человеческого ТПА немутантного типа. Другой вариант ТПА гликозилирован в позиции 105 последовательности аминокислот человеческого ТПА немутантного типа. Еще один вариант ТПА гликозилирован в позициях 103-107 через -связь. Другой вариант ТПА гликозилирован в позициях 103-105 через О-связь. Один из вариантов ТПА содержит аспарагин как часть или трипептидной последовательности, где Х - любая аминокислота, за исключением пролина, в позиции 103 последовательности аминокислот человеческого ТПА немутантного типа. Предпочтительно вариант ТПА содержит аспарагин в позиции 103, триптофан в позиции 104 и серин в позиции 105 аминокислотной последовательности человеческого ТПА немутантного типа. Еще один вариант ТПА содержит аспарагин как часть или трипептидной последовательности, где Х - любая аминокислота, за исключением пролина, в позиции 105 последовательности аминокислот человеческого ТПА немутантного типа. Предпочтительно вариант ТПА содержит аспарагин в позиции 105, треонин в позиции 106 и серин в позиции 107 последовательности аминокислот человеческого ТПА немутантного типа. Один из вариантов ТПА содержит другую аминокислоту, чем аспарагин, в позиции 117 последовательности аминокислот человеческого ТПА немутантного типа. Предпочтительно вариант ТПА содержит глутамин в позиции 117 последовательности аминокислотчеловеческого ТПА немутантного типа. Другой предпочтительный вариант ТПА дополнительно содержит аспарагин как часть или трипептидной последовательности, где Х - любая аминокислота, за исключением пролина, замещенная в позиции 103 последовательности аминокислот человеческого ТПА немутантного типа. Еще один вариант ТПА дополнительно содержит аспарагин как часть или трипептидной последовательности, где Х - любая аминокислота, за исключением пролина, в позиции 105 последовательности аминокислот человеческого ТПА немутантного типа. Предпочтительно вариант ТПА дополнительно содержит серин в позиции 107 последовательности аминокислот человеческого ТПА немутантного типа. Один из вариантов ТПА имеет повышенную фибриноспецифичность по сравнению с человеческим ТПА немутантного типа. 4094 1 В предпочтительном варианте ТПА указанная фибриноспецифичность достигается путем иного изменения, чем замещение аспарагиновой кислоты аргинином в позиции 299, в пределах аминокислотной области 296-302 или 274-277 последовательности аминокислот человеческого ТПА немутантного типа. В другом варианте ТПА указанное изменение находится в области 296-300 последовательности аминокислот человеческого ТПА немутантного типа. Предпочтительно вариант ТПА указанное изменение находится в одной или более позициях 296, 297, 298 и 299 последовательности аминокислот человеческого ТПА немутантного типа, при этом указанное изменение является замещением аланина каждой из аминокислот лизином, гистидином, аргинином в позициях 296,297, 298 и 299 последовательности амимнокислот человеческого ТПА немутантного типа. Один из вариантов ТПА содержит аспарагин как часть или трипептидной последовательности в позициях 103 и аминокислоту, иную, чем аспарагин, в позиции 117 последовательности аминокислот человеческого ТПА немутантного типа. Предпочтительный вариант ТПА содержит аспарагин в позиции 103, триптофан в позиции 104, серин в позиции 105 и глутамин в позиции 117 последовательности аминокислот человеческого ТПА немутантного типа. Другой вариант ТПА содержит аспарагин как часть или трипептидной последовательности в позиции 105 и аминокислоту, иную, чем аспарагин, в позиции 117 последовательности аминокислот человеческого ТПА немутантного типа. Предпочтительно вариант ТПА содержит аспарагин в позиции 105, треонин в позиции 106, серин в позиции 107 и глутамин в позиции 117 последовательности аминокислот человеческого ТПА немутантного типа. Еще один вариант ТПА выбран из группы, состоящей из вариантов человеческого ТПА, содержащих следующие мутации 103, 117 105, 107, 117 103, 117, К 296 А, Н 297 А, 298, 299 105, 107, 117, К 296 А, Н 297 А, 298, 299 103, 117, 298, 299 105, 107, 117,298, 299 103, 117, 296, 297, 301 105, 107, 117, 296, 297, 301 103,117, 274, 275, 276, К 277 Т 105, 107, 117, 274, 275, 276, К 277 Т, гдеобозначает любую аминокислоту, за исключением аспарагина . Другим объектом изобретения являются последовательности ДНК, кодирующие вариант ТПА, содержащие нуклеотиды,,и. Еще одним объектом изобретения является состав для лечения сосудистого нарушения или заболевания,содержащий терапевтически активное вещество и фармацевтически приемлемый носитель, причем в качестве терапевтически активного вещества он содержит вариант ТПА в эффективном количестве. Объектом изобретения является также способ лечения сосудистого нарушения или заболевания у млекопитающих, при котором млекопитающему вводят эффективное количество вышеуказанного состава. Еще одним объектом изобретения является состав для предотвращения отложения фибрина или образования или преобразования адгезии, содержащий терапевтически активное вещество и фармацевтически приемлемый носитель, при этом в качестве терапевтически активного вещества он содержит вариант ТПА в эффективном количестве. Объектом изобретения является также способ лечения млекопитающего для предотвращения отложения фибрина или образования или преобразования адгезии, при котором в сайт потенциального образования фибрина или адгезии у млекопитающего вводят эффективное количества вышеуказанного состава. Еще одним объектом изобретения является способ восстановления свойства связывания фибрина с человеческим ТПА немутантного типа, при котором ТПА редуцируют в результате гликозилирования в любой из аминокислотных позиций 103-105 и удаляют функциональную углеводную структуру в аминокислотной позиции 117. Исследования продемонстрировали, что потеря ТПА способности связывания с фибрином, возникающая в результате изменения его первичной функции, которое должно улучшить его фармакокинетические свойства (уменьшение очистки плазмы, расширение циркулирующего периода полураспада) ТПА немутантного типа, восстановливается путем дополнительного изменения без компромисса с более медленной скоростью очистки или достижением расширенного циркулирующего периода полураспада. Экспериментально доказано, что (фибринолитическая) активность лизиса тромба таких вариантов ТПАзначительно повышается по сравнению с активностью человеческого ТПА немутантного типа, в частности, если изменения способа гликозилирования ТПА сопровождаются дополнительными специфическими изменениями в протеазном домене ТПА. В результате образуются молекулы с повышенной фибринолитической активностью по сравнению с ТПА немутантного типа, которые также способны к более быстрому лизису тромбов в плазме, чем у ТПА немутантного типа, и, кроме того, имеют повышенную специфичность связывания с фибрином. Было обнаружено, что потеря способности связывания с фибрином, наблюдаемая для вариантов ТПА с медленной очисткой, имеющих сайт повышенного гликозилирования в позициях 103-105 аминокислот человеческого ТПА немутантного типа, восстанавливается путем удаления углеводной структуры в позиции аминокислоты 117. Эти изменения в сочетании расширяют циркулирующий период полураспада без ком 7 4094 1 промисса со способностью к связыванию с фибрином варианта ТПА. Фибринолитическая активность(лизис тромбов на единичную дозу) ТПА немутантного типа также повышается, когда молекулу подвергают дополнительным изменениям, улучшающим специфичность связывания с фибрином. Изобретение иллюстрируется на чертежах, на которых на фиг. 1 и 2 согласно настоящему изобретению показано влияние мутаций на связывание с фибрином результирующих вариантов ТПА. Потеря способности связывания с фибрином (фибриноспецифичность связывания) из-за повышенного гликозилирования в позиции 103 аминокислоты человеческого ТПА немутантного типа может быть восстановлена путем удаления углеводной структуры в позиции 117 молекулы человеческого ТПА немутантного типа. Фиг. 3, 4, 5 и 6 показывают результаты анализа лизиса тромбов, выполненного на модели тромболиза артерио-венозного анастомоза у кролика. Термины ТПА, человеческий ТПА и человеческий тканевый плазминогенный активатор относятся к человеческому внешнему (тканевого типа) плазминогенному активатору, имеющему фибринолитическую активность, который имеет структуру с 5 доменами (, фактора роста, -1, -2 и протеазным доменами), но тем не менее может иметь меньше доменов или некоторые из этих доменов могут повторяться, если он еще функционирует как тромболитический агент. Как минимум, ТПА состоит из протеазного домена, который способен преобразовать плазминоген в плазмин, и области терминали, которая, полагают,по крайней мере частично ответственна за связывание фибрина. Таким образом, эти термины обозначают полипептиды, содержащие эти функциональные домены как часть последовательности аминокислот полипептида. Биологически активные формы ТПА могут быть получены системами культивирования рекомбинантных клеток в формах, содержащих две функциональные области молекулы и любые другие части ТПА,в других отношениях свойственных природному источнику ТПА. Понятно, что существуют нативные аллельные вариации и могут иметь место среди индивидуумов, о чем свидетельствуют одно или более различий в последовательности аминокислот ТПА каждого индивидуума. Термины ТПА немутантного типа, нативный ТПА, человеческий ТПА немутантного типа и нативный человеческий ТПА относятся к человеческому ТПА с нативной последовательностью, т.е. к тому, который закодирован последовательностью к ДНК, изложенной в патенте США 4 766 075, выданном 23 августа 1988 г. Номера сайтов или позиций аминокислот в молекуле ТПА обозначаются в соответствии с патентом США 4 766 075. ТПА может быть выделен из любого природного источника. Кроме того, ТПА может быть получен из любой рекомбинантной системы экспрессии, включая, например, яичники китайского хомяка (клетки СНО) или клетки линии 293 почки человеческого эмбриона. Термины связывание фибрина и фибриноспецифичность связывания относятся к способности молекулы ТПА связывать фибриновые тромбы в стандартных пробах по связыванию с фибрином, таких как способ, описанный., . . . 257, 2920-2925 (1982), или в ее модифицированной версии, раскрытой в примере 2. Термины биологическая активность ТПА, биологически активный, активность и активный относятся к способности молекулы ТПА преобразовывать плазминоген в плазмин, как измерено в пробе -2251 в присутствии тромба в плазме или в присутствии фибрина, в пробе -2288, в пробе лизиса тромба в плазме или других соответствующих пробах. Проба (пробы) могут проводиться в присутствии или отсутствии потенциальных модуляторов активности, таких как фибрин, фибриноген, плазма и/или тромбы в плазме. Выражения фибринолитическая активность, тромболитическая активность и активность лизиса тромбов используются равнозначно и относятся к способности молекулы ТПА растворять тромб, полученный из очищенного фибрина или из плазмы, используя любую пробу лизиса тромба, известную из уровня техники, такую, как проба лизиса очищенного тромба., .168, 428-435(1988) и ее модифицированная форма, описанная , .,1991, выше. Выражения специфическая фибринолитическая активность, специфическая тромболитическая активность и специфическая активность лизиса тромба относятся к лизису тромба при единичном устойчивом состоянии уровня плазмы, как определяется любой пробой лизиса тромба, известной из уровня техники, такой, как упоминалась выше. Выражения фибринолитическая активность, тромболитическая активностьи активность лизиса тромбаиспользуются равнозначно и относятся к лизису тромба на единичную дозу ТПА. Фибринолитическая активностьопределяется в любой принятой живой модели пробы лизиса тромба, включая модель легочной эмболии хомяка (, .а 1., 1991, выше) и модель тромбоза яремной вены кролика .., . 74, 368 (1983). В частности, предпочтительный вариант последней модели описан в нижеследующем примере. Выражение в значительной степени остаточное связывание с фибрином (специфичность) (по сравнению с человеческим ТПА немутантного типа) и его грамматические варианты, используемые здесь, обозначают, что способность связывания с фибрином (кажущееся значение К) молекулы варианта ТПА находится в пределах около двух порядков способности к связыванию фибрина (значение К) для человеческого ТПА немутантного типа, как определено в той же самой пробе. Выражение в значительной степени повышенное связывание фибрина относится к возрастанию более чем на 4 порядка способности связывания с фибрином 8(кажущееся значение ) варианта ТПА, полученного включением дополнительной мутации или набором мутаций. Термин повышеннаяфибролитическая активность по сравнению с ТПА немутантного типа относится к сравнительному лизису тромба, достигаемому при введении варианта ТПА в дозе,равной примерно одной трети или менее дозы ТПА немутантного типа. Термины скорость очистки и очистка относятся к скорости, с которой молекула ТПА удаляется из кровяного потока. Очистка (скорость) измеряется относительно нативного ТПА, при этом пониженная очистка (скорость) указывает, что вариант ТПА очищается медленнее, чем нативный ТПА, а повышенная очистка (скорость) указывает, что вариант ТПА очищается быстрее, чем нативный ТПА. Выражение фибриноспецифичность относится к активности мутанта, который проявляет более высокое соотношение фибрино-зависимой специфической активности к фибриногенно-зависимой специфической активности в пробе 2251, чем человеческий рТПА немутантного типа. Это соотношение преимущественно составляет, по крайней мере, 15. Выражение плазменная тромбовая специфичность относится к активности мутанта, который проявляет более высокое соотношение плазменной тромбово-зависимой специфической активности к плазменнозависимой специфической активности в пробе 2251, чем рТПА немутантного типа. Это соотношение преимущественно составляет, по крайней мере, 15. Термины зимоген, зимогенный и зимогенная активность используются здесь, как указано в заявке 90/02798, опубликованной 22 марта 1990 г. В соответствии с этим определением не является необходимым полное отсутствие ферментативной активности. Выражение удаление функциональной углеводной структуры в позиции 117 аминокислот человеческого ТПА немутантного типа обозначает полное удаление углевода в остатке аминокислоты 117, как в случае,когда сигнал гликозилирования отменяется мутагенезом, направленным на сайт, как описано ниже, или значительное удаление, как в случае обработки эндогликозидазой, которая может, например, оставить нетронутым остаток -ацетилглюкозамина, связанный с 117. Термины аминокислота и аминокислоты относятся ко всем нативно присутствующим аминокислотам. Это определение подразумевает включение норлейцина, орнитина и гомоцистеина. Аминокислоты обозначаются или одной буквой, или тремя буквамиаспарагиновая кислотаТ треонинсеринЕ глутаминовая кислотаР пролинглицин А 1 а А аланинС цистеинвалинМ метионин еизолейцинлейцин ТутирозинфенилаланингистидинК лизинаргининтриптофанглутаминаспарагин Эти аминокислоты могут классифицироваться в соответствии с химическим составом и свойствами их боковых цепей. Они широко классифицируются по двум группам насыщенные и ненасыщенные. Каждая из этих групп делится на подгруппы для более тщательной классификации аминокислот 1. Насыщенные аминокислоты Кислотные остатки аспарагиновая кислота, глутаминовая кислота. Основные остатки лизин, аргинин, гистидин.. Ненасыщенные аминокислоты Гидрофильные остатки серии, треонин, аспарагин, глутамин. Алифатические остатки глицин, аланин, валин, лейцин, изолейцин. Неполярные остатки цистеин, метионин, пролин Ароматические остатки фенилаланин, тирозин, триптофан. Термины изменение, изменение последовательности аминокислот и вариант последовательности аминокислот относятся к молекулам ТПА с некоторыми различиями в последовательности их аминокислот по сравнению с нативным ТПА. Обычно варианты обладают, по меньшей мере, 80-ной гомологией с теми 9 4094 1 доменами нативного ТПА, которые остаются в их структуре, и преимущественно они будут составлять, по крайней мере, 90-ную гомологию с такими доменами. Варианты гликозилирования ТПА, относящиеся к сфере этого изобретения, могут также по выбору включать замещение, делецию и/или вставки дополнительно к тем, которые являются результатом специфических изменений в способе гликозилирования ТПА. Замещенными вариантами ТПА являются такие, в которых удален, по меньшей мере, один остаток аминокислоты в последовательности нативного ТПА и на его место в ту же самую позицию вставлена другая аминокислота. Замещения могут быть одиночными, когда замещена только одна аминокислота в молекуле,или они могут быть многократными, когда две или более аминокислоты замещены в одной и той же молекуле. Значительные изменения активности молекулы ТПА могут быть получены путем замещения аминокислотой с боковой цепью, которая существенно отличается по насыщенности и/или структуре от нативной аминокислоты. Предполагается, что этот тип замещения влияет на структуру полипептидной основы и/или насыщенность или гидрофобность молекулы в области замещения. Умеренные изменения активности молекулы ТПА могут ожидаться при замещении аминокислотой с боковой цепью, которая подобна по насыщенности и/или структуре аминокислоте нативной молекулы. Предполагается, что этот тип замещения, относящийся к консервативному замещению не будет существенно изменять ни структуру полипептидной основы, ни насыщенность или гидрофобность молекулы в области замещения. Варианты вставки ТПА - это варианты с одной или более аминокислотами, вставленными непосредственно смежно с аминокислотой в конкретной позиции в молекуле нативного ТПА. Непосредственно смежная связь с аминокислотой обозначает связь или с -карбокси- или с -амино-функциональными группой аминокислоты. Вставкой может быть одна или более аминокислота. Обычно вставка будет содержать одну или две консервативных аминокислоты. Аминокислоты, подобные по насыщенности и/или структуре аминокислотам, смежным с местом вставки, определяются как консервативные. Альтернативно настоящее изобретение включает вставку аминокислоты с насыщенностью и/или структурой, которые в значительной степени отличаются от аминокислот, смежных с местом вставки. Варианты с делецией - это варианты с удаленными одной или более аминокислотами в молекуле нативного ТПА. Обычно в вариантах с делецией будут удалены одна или более аминокислот в конкретной области молекулы ТПА. Ниже раскрывается система обозначений, применяемая здесь для описания вариантов последовательности аминокислот ТПА,. Положение конкретной аминокислоты в полипептидной цепочке ТПА обозначается номером. Номер относится к позиции аминокислоты в последовательности аминокислот полипептида сформированного человеческого ТПА немутантного типа (патент США 4 766 075, выданный 23 августа 1988 г.). В настоящем применении варианты остатков, подобным образом расположенных в ТПА, обозначаются теми же номерами в результате удалений или вставок в молекулу. Например, это может иметь место в вариантах с делецией, направленной на сайт, или со вставкой. Аминокислоты обозначаются с использованием однобуквенного кода. Замещенные аминокислоты обозначены путем указания аминокислоты немутантного типа слева от номера, обозначающего позицию в полипептидной цепочке этой аминокислоты, и указания замещенной аминокислоты справа от номера. Например, замена аминокислоты треонина (Т) аспарагиномв позиции аминокислоты 103 молекулы человеческого ТПА немутантного типа дает вариант ТПА, обозначенный 103 ТПА. Аналогично, вариант ТПА, полученный дополнительным замещением глутаминааспарагиномв аминокислотной позиции 117 молекулы человеческого ТПА немутантного типа обозначается 103, М 117 ТПА. Варианты с делецией обозначаются путем указания остатка аминокислоты и позиции в одном конце делеции, включая и размещение греческой буквы дельтаслева от указанных аминокислот. Например, вариант ТПА, содержащий делецию аминокислот 296-299, будет обозначаться как 296-297-298-299 ТПА, где К, Н иобозначают соответственно аминокислоты - лизин, гистидин и аргинин. Делеция одной аминокислоты, например, К 296, будет обозначаться как К 296. Вставочные варианты ТПА обозначаются с использованием квадратных скобоквокруг вставленных аминокислот, а расположение вставки отмечается указанием позиции аминокислоты с одной стороны вставки. Например, вставка аминокислоты аланина(А) между глутаминовой кислотой в позиции 94 и аспарагиновой кислотой в позиции 95 будет обозначаться как 9495. Для облегчения чтения запятая (,) используется для разделения многократных мутаций,которые имеют место в отдельной молекуле, а точка с запятойиспользуется для разделения молекул отдельного варианта ТПА, который был составлен там, где перечислены вместе несколько молекул варианта ТПА. Термины кодирование последовательности ДНК, кодирование ДНК и кодирование нуклеиновой кислоты относятся к порядку или последовательности дезоксирибонуклеотидов вдоль цепочки дезоксирибонуклеиновой кислоты. Порядок этих деоксирибонуклеотидов определяет порядок аминокислот вдоль полипептидной цепочки. Таким образом, последовательность ДНК кодирует последовательность аминокислот. 4094 1 Термины воспроизводимый вектор экспрессии и вектор экспресии относятся к части ДНК, обычно с двойной цепочкой, которая может быть вставлена в часть чужеродной ДНК. Чужеродная ДНК определяется как гетерогенная ДНК, которая является ДНК, не находящейся нативно в клетке-реципиенте. Вектор используется для переноса чужеродной или гетерогенной ДНК в соответствующую клетку-реципиент. Находящийся в клетке-реципиенте вектор может быть реплицирован независимо от основной хромосомной ДНК, и могут быть получены несколько копий вектора и вставленные (чужеродные) ДНК. Кроме того, вектор содержит необходимые элементы, которые обеспечивают перенос чужеродной ДНК в полипептид. Таким образом могут быть быстро синтезированы многие молекулы полипептида, закодированные чужеродной ДНК. Нуклеиновая кислота оперативно связана, когда она находится в функциональном соотношении с последовательностью другой нуклеиновой кислоты. Например, ДНК для предварительной последовательности или секреторной лидерной последовательности оперативно связана с ДНК, кодирующей полипептид, если она закодирована как белок, который участвует в секреции полипептида промотор или усилитель оперативно связан с кодирующей последовательностью, если он влияет на транскрипцию последовательности или сайт рибосомной связи оперативно связан с кодирующей последовательностью, если он расположен для обеспечения трансляции. В общем, оперативно связан означает, что связанные последовательности ДНК являются смежными и в случае секреторной лидерной последовательности проводника - смежными также в фазе считывания. Однако усилители не должны быть смежными. Связь выполняется связыванием в удобных рестрикционных сайтах. Если таких сайтов не существует, то используются синтетические олигонуклеотидные адаптеры или линкеры в соответствии с обычной практикой. В контексте настоящего изобретения выражения клетка, линия клетки и культура клетки используются равнозначно, и все эти обозначения включают потомков. Термины трансформированная клетка-реципиент, трансформант и трансформированный относятся к вводу ДНК в клетку. Клетка называется основной клеткой-реципиентом. Введенная ДНК обычно существует в форме вектора, содержащего введенную часть ДНК. Введенная последовательность ДНК может состоять из тех же частей, что и клетка-реципиент, или из частей, отличных от клетки-реципиента, или она может быть гибридной последовательностью ДНК, содержащей некоторую чужеродную и некоторую гомологическую ДНК. Слова трансформанты и трансформированные клетки-реципиенты включают первичный объект- клетку и культуры, полученные из нее, независимо от количества перемещений. Понятно, что все потомки могут быть не точно идентичными по содержанию ДНК из-за преднамеренных или ненамеренных мутаций. Включаются потомки мутанты, имеющие те же самые функциональные или биологические свойства, что и отбираемые в первоначально трансформированной клетке. Плазмиды обозначены индексом р, следующим внизу за буквенно-цифровым обозначением. Начальные плазмиды, используемые в этом изобретении, поставляют коммерчески в неограниченном неограниченном количестве, или могут быть получены из поставляемых плазмид с использованием опубликованных процедур. Кроме того, для обычного специалиста могут быть очевидными другие эквивалентные плазмиды,известные в этой области. 2. Общие методы А. Выбор вариантов Гликозилирование полипептидов обычно является -связанным или О-связанным. -связь обычно относится к прикреплению углеводной половины к боковой цепочке аспарагинового остатка. Трипептидные последовательности аспарагин-Х-серин и аспарагин-Х-треонин, в которых Х является любой аминокислотой,за исключением пролина, являются признанными последовательностями для ферментативного прикрепления углеводной половины к аспарагиновой боковой цепочке. Гликозилирование через О-связь относится к прикреплению одного из сахаров -ацетилгалактозамина, галактозы или ксилозы к гидроксиаминокислоте, чаще всего к серину или треонину, хотя 5-гидроксипролин или 5-гидроксилизин также могут быть вовлечены в гликозилирование через О-связь. Способы гликозилирования для протеинов, вырабатываемых млекопитающими, подробно описаны в,, , , ., 2 , .4,, , 1984, 271-315. В этой главе обсуждаются связанные с аспарагином олигосахариды,включая их подразделения, по меньшей мере, на 3 группы, отнесенные к сложной структуре, структуре с высоким содержанием маннозы и гибридной структуре, а также олигосахариды, связанные посредством гликозилирования. Способы гликозилирования для белков, полученные при помощи дрожжей, подробно изложены, . . . 906(1), 915-944 (1987). Типичные схемы гликозилирования белков через -связь, полученные из клеток млекопитающих и дрожжей, сравниваются в фиг. 1 в заявке 89/11531, опубликованной 30 ноября 1989 г. Предварительные исследования углеводных структур нативного ТПА, очищенного от клеток меланомы(меланомный ТПА, мТПА), продемонстрировали присутствие олигосахаридов с высоким содержанием маннозы в позиции -117 с олигосахаридами сложного типа в позициях 184 и 448 (.,11. 170, 69-75 (1987). ТПА типа 1 имеет все 3 позиции с -связью, замещенные углеводом, в то время, как ТПА типане имеют углевода, связанного с позицией 184, соответственно эта позиция гликозилирована примерно только в 50 молекул нативного ТПА. В одном подробном отчете.,28, 7644-7662 (1989) подтверждены эти распределения, и кроме того, обнаружено, что было сульфатировано 30 олигосахаридов. Также были определены олигосахариды для рекомбинантного ТПА немутантного типа, очищенного от транфектированных клеток яичника китайского хомяка (СНО) (рТПА, или ТПА),.,, 264, 1410014111 (1989), которые, как было обнаружено, имеют подобный способ гликозилирования олигосахариды с высоким содержанием маннозы в 117, сложные олигосахариды в 184 и 448, с сайтом 184, в котором гликозилировано только 50 молекул. Варианты этого изобретения должны содержать, по крайней мере, одну последовательность аминокислот, имеющую потенциал для осуществления гликозилирования через - и Освязи в любой из позиций аминокислот 103, 104 и 105 человеческого ТПА немутантного типа. Если рассматривать гликозилирование с -связью, сайт гликозилирования в варианте является трипептидной последовательностью формулы аспарагин-Х-серин или аспарагин-Х-треонин, в которой аспарагин является акцептором, а Х является одной из 20 генетически закодированных аминокислот, за исключением пролина, который, как известно, препятствует гликозилированию. См. , , в, .,, 1980, .35 , . . 40, 17(1974) и , в, , .,, , 1973,1-15. Варианты последовательностей аминокислот здесь модифицированы или вставкой подходящих аминокислот(ы) в соответствующий сайт(ы) для обеспечения гликозилирования (как при добавлении аспарагин-Х-серин/треонин трипептида после позиции 103, чтобы сделать плоскую петлю больше и более открытой) или замещением аминокислот(ы) в соответствующих сайтах подходящими аминокислотами для осуществления гликозилирования. Если должно быть использовано гликозилирование с О-связью, О-гликозилированная связь имеет место в клетках животных между -ацетилгалактозамином, галактозой или ксилозой и одной из нескольких гидроксиаминокислот, чаще всего серином или треонином, но в некоторых случаях также с 5 гидроксипролиновым или 5-гидроксилизиновым остатком, размещенным в соответствующей области молекулы. В одном из предпочтительных вариантов выполнения гликозилирования с -связью в сайте добавляют аминокислоты в позициях 103-105 нативного человеческого ТПА. Для гликозилирования через Освязь заменяют одну или более аминокислот в этих областях или добавляют серин, треонин или остаток 5 гидроксилизина. Такие варианты ТПА описаны в 89/11531, выше. Варианты ТПА по настоящему изобретению дополнительно отличаются отсутствием функциональной углеводной структуры в остатке аминокислоты 117. В соответствии с предпочтительным вариантом выполнения функциональная углеводная структура сохраняется во всех других позициях аминокислот, гликозилированных в молекуле нативного ТПА. Такое селективное удаление функциональной углеводной структуры в позиции 117 предпочтительно достигается замещением аминокислот, по крайней мере, для одного остатка в сигнале гликозилирования в позициях 117-119 последовательности аминокислот человеческого ТПА немутантного типа. В особенно предпочтительном варианте аспарагинв позиции аминокислоты 117 замещается другой аминокислотой, предпочтительным заместителем является глутамин(см. например, Европейскую патентную заявку 238 304, выше, и в 89/04368). В предпочтительных вариантах настоящего изобретения аспарагин замещен треонином в позиции 103 или серином в позиции 105 в сочетании с замещением серина аланином в позиции 107 нативного человеческого ТПА, а аспарагинв позиции 117 замещается любой другой аминокислотой, предпочтительным заместителем является глутамин . Плазминогенные активаторы, в дополнение к предшествующим модификациям в способе гликозилирования нативного ТПА, по выбору содержат также замещения, делеции или вставки остатков в других областях последовательности нативного ТПА для улучшения некоторых свойств молекулы. Такие модификации хорошо известны в этой области. Общий обзор плазминогенных активаторов и их производных второго поколения можно найти в ,, 1 449-458 (1987) и , , ., .. 66 (1) 88-110 (1991). Другие обзоры вариантов ТПА изложены., , 2 123132 (1988),а 1., в, . 23,12 (1988) и, 1990, выше. Например, средством дальнейшего улучшения скорости очистки и/или периода полураспада является удаление части или всехдоменов и/или домена фактора роста из вариантов ТПА по настоящему изобретению. Альтернативно или дополнительно варианты ТПА могут оказать сопротивление протеолитическому расщеплению в аминокислотах или около аминокислот 275 и 276 и/или имеющих модификации аминокислотах в сайте предполагаемой связи с лизином домена -2 ТПА. 12 4094 1 Считалось, что особенно желательно обеспечить варианты ТПА, которые, кроме их более медленной очистки являются более фибриноспецифичными, чем ТПА немутантного типа. Такие варианты ТПА будут действовать предпочтительно в сайте фибринового тромба по сравнению с модифицированным ТПА, и, следовательно, как ожидается, вызовут пониженные побочные эффекты, связанные с деятельностью циркулирующего плазминогена, например, менее сильные и менее частые осложнения, связанные с кровотечением. Фибриноспецифичность вариантов ТПА по настоящему изобретению может быть улучшена какимнибудь дополнительным изменением, известным из уровня техники. Для улучшения фибриноспецифичности вариант ТПА по настоящему изобретению может, например,дальше подвергаться мутации в позициях 296-302 аминокислот, преимущественно в позициях 296-299 домена сериновой протеазы. В предпочтительном варианте каждая из аминокислот лизин (К), гистидин (Н), аргининв позициях 296-299 ТПА немутантного типа замещена аланином. В следующем предпочтительном варианте оба аргинина в позициях 298 и 299 заменены глутаминовой кислотой. В другом предпочтительном варианте лизин (К), гистидин (Н) и пролин (Р) в позициях 296, 297 и 301 аминокислот ТПА немутантного типа дополнительно замещены соответственно глутамином , аспарагиноми серином . В следующем предпочтительном варианте фибриноспецифичность вариантов ТПА по настоящему изобретению улучшается путем замещения фенилаланина , аргинина , изолейцинаи лизина (К) в позициях 274,275, 276 и 277 в последовательности аминокислот человеческого ТПА немутантного типа соответствующими аминокислотами лейцином , гистидином (Н), сериноми треонином (Т). Последнее изменение вызывает потерю сайта плазменного расщепления следовательно, варианты находятся в основном в одноцепочной форме. Кроме того, молекулы по этому изобретению могут быть замещены или могут содержать делеции в некоторых позициях для придания дополнительных желательных свойств, включая зимогенный характер. Эти позиции человеческого ТПА включают, например, замещение аланина лизином, гистидином и глутаминовой кислотой в позициях 416-418 соответственно и замещение аланина глутаминовой кислотой в позициях 426,427, 429 и 430 соответственно, как изложено, например, в 90/02798, опубликованной 22 марта 1990 г. Примерами подходящих многократных мутантов являются 103, 117 ТПА 105, 107, 117 ТПА 103, 117,(296-299)АААА ТПА 105, 107, 117, (296-299) ТПА 103, 117, 298, 299 ТПА 105, 107, 117, 298, 299 ТПА 103, 117, 296,297, 301 ТПА 105, 107, 117, 296, 297, 301 ТПА 103, 117, (274-277) ТПА 105, 107, 117, (274-277) ТПА, гдеобозначает любую из нативно присутствующих 20 аминокислот, за исключением аспарагина . Особенно предпочтительными являются варианты ТПА, в которых в позиции 117 аспарагинзамещен глутамином . В. Конструкция вариантов Проведение повышенного гликозилирования среди нативных молекул ТПА и удаление функциональной углеводной структуры в аминокислотном остатке 117 могут быть выполнены способами, известными из уровня техники. Химическая и ферментативная связь гликозидов с протеинами может быть осуществлена с использованием множества активированных групп, например, как описаноив. . . . 259-306 (1981). Преимущества техники химической связи заключаются в том, что она относительно проста и не нуждается в сложной ферментативной обработке, которая требуется для нативного гликозилирования с Ои - связями. В зависимости от используемого вида связи сахар(а) может (могут) быть присоединен(ы) к (а) аргинину и гистидину, (б) к свободным карбоксильным группам, таким, как группы глутаминовой кислоты и аспарагиновой кислоты (с) к свободным сульфгидриловым группам, таким как группы цистеина, (д) к свободным гидроксильным группам, таким как группы серина, треонина или гидроксипролина, (е) к ароматическим остаткам, таким как остатки фенилаланина, тирозина или триптофана или (ж) к амидной группе глутамина. Эти способы описаны в 87/05330 (опубликована 11 сентября 1987 г.). Например, углеводная структура в аминокислотной позиции 117 нативного человеческого ТПА может быть в значительной степени удалена при использовании эндогликосидазы, такой как эндогликосидаза Н(-). - способна только на (частичное) удаление олигосахаридов с высоким содержанием маннозы и гибридных олигосахаридов. Соответственно при соответствующих условиях - способна (в значительной степени) удалять углеводную структуру с высоким содержанием маннозы в аминокислотном остатке 117 нативного ТПА без функционального влияния на сложные структуры в аминокислотных остатках 184 и 448. Эта обработка выполняется с использованием известной по существу технологии, например, в соответствии с методом., 249, 811 (1974),., . . 141, 515(1984) и., . 23, 6191 (1984). Варианты ТПА по настоящему изобретению в основном создаются путем мутации последовательности ДНК, которая кодирует ТПА немутантного типа, и экспресси мутированной последовательности ДНК в соответствующую клетку-реципиент. Модификация с изменением или вставкой соответствующих аминокислот(ы) в нативную молекулу для получения желательных вариаций последовательности выполняется любыми способами, известными из 13 4094 1 уровня техники, такими как, например, направленный на сайт мутагенез или связывание соответствующей последовательности в ДНК, кодирующую релевантный белок, как описано ниже. В то время, как любая технология, известная из уровня техники, может быть использована для выполнения мутагенеза, направленного на сайт, например, как описано в.,,,,(1989), направленный на олигонуклеотид мутагенез является предпочтительным способом приготовления вариантов ТПА по настоящему изобретению. Этот способ, который хорошо известен из уровня техники (., , 2 183 (1983),., выше), является наиболее подходящим для синтеза замещенных вариантов, он может быть также использован для вариантов, которые обычно получает делецией и вставкой. Олигонуклеотиды легко синтезируются с использованием технологии, хорошо известной из уровня техники, такой как описано., (. , 755765 1978). Мутанты с более чем одной замещенными аминокислотами, могут быть получены одним из нескольких способов. Если аминокислоты размещены близко друг к другу в полипептидной цепи, они могут одновременно мутировать при использовании одного олигонуклеотида, который кодирует все желательные замещения аминокислот. Однако, если аминокислоты размещены на некотором расстоянии друг от друга (например, разделены более чем 10 аминокислотами), то труднее получить один олигонуклеотид, который кодирует все желательные изменения. Вместо этого может быть использован один из двух альтернативных способов. В первом способе получают отдельный олигонуклеотид для каждой аминокислоты, которая должна быть замещена. Затем олигонуклеотиды одновременно отжигают до одноцепочной модели ДНК, а вторая цепочка ДНК, которая синтезируется из матрицы, будет кодировать все желательные замещения аминокислот. Альтернативный способ включает два или более цикла мутагенеза для получения желательного мутанта. Другой способ получения мутаций в последовательности ДНК, кодирующей ТПА немутантного типа или молекулу варианта, известный из уровня техники, например, для получения нового сайта гликозилирования в соответствии с настоящим изобретением, включает расщепление последовательности ДНК, кодирующей начальную молекулу ТПА в соответствующей позиции путем сжигания с рестрикционными ферментами,восстановления надлежащим образом расщепленной ДНК, синтеза олигонуклеотида, кодирующего желательную последовательность аминокислот для гликозилирования и фланкирующих областей, таких как полинкеры с тупыми концами (или, вместо полилинкеров также используется сжигание синтетического олигонуклеотида с рестрикционными ферментами для расщепления ТПА, кодирующего ДНК, получая этим концы, способные к сцеплению), и ДНК с остатком ТПА, кодирующего структурный ген.-мутагенез также пригоден для получения вариантов ТПА по настоящему изобретению как, например, изложено в патенте США 4 683 195, выданном 28 июля 1987 г., и в,., .-,2,15, 1991. По настоящему изобретению, кДНК, кодирующая варианты ТПА, вставляется в реплицируемый вектор для дальнейшего клонирования или экспрессии. Подходящие векторы готовят с использованием стандартных способов получения рекомбинантной ДНК. Для получения желательных векторов изолированные плазмиды и фрагменты ДНК расщепляют, стыкуют и сшивают вместе в специфическом порядке. После сшивки вектор с вставленным чужеродным геном сейчас же трансформируют в соответствующую клетку-реципиент Селекцию трансформированных клеток осуществляют, используя рост на антибиотике,обычно тетрациклинеили ампицилине (ар), к которым они устойчивы благодаря наличию в векторе генов устойчивости ки/или ар. Если связующая смесь трансформирована в эукариотную клеткуреципиент, трансформированные клетки могут быть выбраны системой /. Трансформированные клетки выращивают в культуре, и затем изолируют плазмиду ДНК (плазмида относится к вектору, связанному с чужеродным интересующим геном). Эту плазмиду ДНК затем анализируют путем рестрикционного картирования и/или секвенирования ДНК. Секвенирование ДНК обычно выполняют или методом.,.,9 309 (1981) или методом.,, 65499 (1980). Согласно этому изобретению прокариоты являются предпочтительными клетками-реципиентами для начальных этапов клонирования. В частности, их используют для быстрого получения больших количеств ДНК, для получения одноцепочных матриц ДНК, используемых для мутагенеза, направленного на сайт, для одновременного скрининга многих мутантов и для секвенирования ДНК у полученных мутантов. Подходящие прокариотные клетки-реципиенты включают штамм 294 . К 12(АТСС 31446), штамм 3110. (27325), . 1776 (31537) и . В однако, также могут быть использованы многие другие штаммы , таких как НВ 101, 101, 522, 538, 539, и многие другие виды и роды прокариот. Они больше предназначены быть иллюстрацией, чем ограничением. Плазмидные векторы, содержащие реплицирующую и контролирующую последовательности, которые получают из видов, совместимых с клеткой-реципиентом, используют с этими клетками-реципиентами. Вектор обычно имеет сайт репликации, маркерные гены, которые обеспечивают фенотипичную селекцию в трансформированных клетках, один или более промоторов и область полилинкера, содержащую несколько сайтов рестрикции для вставки чужеродной ДНК. Плазмиды, обычно используемые для трансформации 14., включают 322, 18, 19, 118, 119 и 13, все они описаны в разделах 1.12-1.20 а 1., выше. Однако, многие другие пригодные векторы также применимы. Эти векторы содержат гены, кодирующие устойчивость к ампицилину и/или тетрациклину, которые дают возможность расти в присутствии этих антибиотиков клеткам, трансформированным с помощью этих векторов. Промоторы, наиболее часто используемые в прокаритных векторах, включают -лактамазу (пенициллиназу) и системы лактозных промоторов (. 375 615 1978., , 1981056 1977., , 281 544 1979 и систему триптофановогопромотора (.,. , 8 4057 1980 Европейская заявка 36 776) и щелочно-фосфатазные системы. В то время,как их наиболее часто применяют, используются и другие микробные промоторы и фрагменты, относящиеся к их нуклеотидным последовательностям, что позволяет квалифицированному специалисту сшить их функционально в плазмидные векторы (см.., , 20269 1980). Для экспрессии вариантов ТПА по настоящему изобретению используют эукариотные микробы (дрожжи) и многоклеточные организмы (клеточные культуры млекопитающих)., или обычные пекарские дрожжи, наиболее часто используют среди низших эукариотных микроорганизмов- реципиентов. Однако, среди множества обычно поставляемых других родов,видов и штаммов, здесь используют такие, как(, , 290140(1981) Европейская патентная заявка 139 383, опубликованная 2 мая 1985 г.(патент США 4 943 529., выше), . ( 98-8, 683, 4574., . .,737(1983), . ( 12424), . ( 16045), . ( 24178) .(Европейская патентная заявка 402226)(Европейская патентная заявка 183 070., . ., 28 265-278 (1988)(Европейская патентная заявка 244 234)(., . . . . , 76 5259-5263 (1979) ,такие какЕвропейская патентная заявка 394 538, опубликована 31 октября 1990 и нитчатые грибы, такие как, например, , ,91/00357, опубликована 10 января 1991 г., и клетки-реципиенты , такие как .., ., 81 1470-1474 (1984) и .,., 4 475-479 (1985). Пригодные промоторные последовательности в дрожжевых векторах включают промоторы для 3- фосфоглицератной киназы (., ., 255 2073 1980) или другие гликолитические ферменты (., ., 7 149 1968., , 17 4900 1978, такие как энолаза, глицеральдигид-З-фосфатдегидрогеназа, гексокиназа, пируватдекарбоксилаза, фосфорфруктокиназа,глюкозо-6-фосфатизомераза, 3-фосфоглицератмутаза, пируваткиназа, триозофосфатизомераза, фосфоглюкозизомераза и глюкокиназа. В конструкциях подходящих плазмид для экспрессии терминальные последовательности, сшитые с этими генами, также сшиты в векторе экспрессии с 3 последовательностью, которую желательно экспрессировать, чтобы обеспечить полиаденилирование мРНК и терминали. Другие промоторы, которые имеют дополнительное преимущество при транскрипции, регулируемой условиями роста, являются областью промотора для алкогольдегидрогеназы 2, изоцитохрома С, кислой фосфатазы, расщепляемых ферментов, связанных с метаболизмом азота, и упоминавшихся выше глицеральдегида-3 фосфатдегидрогеназы и ферментов, отвечающих за утилизацию мальтозы и галактозы. Любой плазмидный вектор, содержащий промотор, сходный с дрожжевым промотором, пригоден для начала репликации и окончания считывания последовательностей. При применении способа согласно этому изобретению могут быть использованы в качестве клетокреципиентов культуры клеток, полученные из многоклеточных организмов. Пригодны культуры клеток как беспозвоночных, так и позвоночных, среди культур клеток позвоночных, в частности, предпочтительны культуры клеток млекопитающих. Примеры пригодных линий клеток включают линиюпочек обезьяны,трансформированную с помощью 40 (-7, АТСС 1651) линию 293 почек человеческого эмбриона (., . . ., 36 59 (1977) клетки почек детеныша хомяка (ВНК, АТСС 10) клетки яичников китайского хомяка (СНО) (, . . . ., 77 4216 1980) клетки Сертоли мыши (ТМ 4, , . ., 23 243 1980) клетки почек обезьньяны (-76, АТСС 70) клетки почек зеленой африканской обезьяны (-76, АТСС 1587) клетки цервикальной карциномы человека (, АТСС 2) клетки почек собаки (, АТСС 34) клетки печени буйволиной крысы ( 3 А, АТСС 1442) клетки легкого человека (138, АТСС 75) клетки печени человека (Нер 2, НВ 8065) клетки опухоли грудной железы мыши (ММТ 060562, АТСС 51) клетки гепатомы крысы (НТС, М 1.54,., . ., 851 1980 и клетки(., . ., 383 44 1982). Векторы экспрессии для этих клеток обычно включают (при необходимости) последовательности ДНК для начала репликации, промотор, размещенный перед геном, который должен быть экспрессирован, сайт связывания с рибосомой, сайт сплайсирования РНК, сайт полиаденилирования и транскрипцию сайта окончания. 4094 1 Промоторы, используемые в векторах экспрессии млекопитающих, часто вирусного происхождения. Эти вирусные промоторы обычно получают из вируса полиомы, аденовируса 2 и наиболее часто встречающегося обезьяньего вируса 40(40). Вирус 40 содержит два промотора, которые называются ранним и поздним промоторами. Эти промоторы особенно пригодны, потому что их оба легко получают из вируса как один из фрагментов ДНК, который также содержит вирусное начало репликации (., , 273 113 1978). Могут также использовать меньшие или большие фрагменты ДНК 40, причем они обеспечивают последовательность приблизительно с 250 п.н. (парами нуклеотидов) от сайтадо сайта В 1, расположенную в начале репликации вируса. Промоторы, которые нативно связаны с чужеродным геном (гомологичные промоторы), могут быть альтернативно использованы при обеспечении совмещения с клеткой-реципиентом, выбранной для трансформации. Начало репликации может быть получено из экзогенного источника, такого как 40 или другой вирус(например, вирус полиомы, аденовирус,(вирус везикулярного стоматита),(вирус папилломы крупного рогатого скота, и вставлено в клонирующий вектор. Альтернативно начало репликации может быть обеспечено хромосомным механизмом репликации клетки-хозяина. Если вектор, содержащий чужеродный ген, интегрирован с хромосомой клетки-реципиента, то последнего часто достаточно. При трансформации клеточных культур синтезируется достаточное количество человеческого ТПА. Однако использование вторичной кодирующей последовательности ДНК может повысить уровни синтеза. Вторичная кодирующая последовательность обычно содержит фермент дигидрофолатредуктазу . Форманемутантного типа обычно подавляется химическим метотрексатом (МТХ). Уровень экспрессиив клетке будет изменяться в зависимости от количества МТХ, добавленного к культивированным клеткам-реципиентам. Дополнительным свойством , которое делает ее особенно пригодной в качестве вторичной последовательности, яляется то, что она может быть использована, как селективный маркер для идентификации трансформированных клеток. Две формыпригодны для использования в качестве вторичных последовательностейнемутантного типа и , устойчивая к МТХ. Тип , используемой в конкретной клетке-реципиенте, зависит от того, будет ли клетка-реципиент лишена(так, что она или синтезирует эндогенно очень низкие уровни , или вообще не синтезирует функциональную ). Линии клеток с нехваткой , такие как линии клеток СНО, описанные(. . . . 77 4216 1980), трансформируют с последовательностями, кодирующиминемутантного типа. После трансформации эти линии клеток с нехваткойэкспрессируют функциональнуюи способны расти в среде культивирования с недостатком питательных веществ, таких, как гипоксантин, глицин и тимидин. Нетрансформированные клетки не выживут в этой среде. Форма , устойчивая к МТХ, может быть использована как средство выбора трансформированных клеток-реципиентов среди тех клеток-реципиентов, которые эндогенно синтезируют нормальные количества функциональной , чувствительной к МТХ. Линия клеток СНО-К 1 (АТСС 61) обладает этими свойствами и, таким образом, является пригодной линией клеток для этой цели. Добавление МТХ к среде культивирования клеток позволит расти только тем клеткам, у которых трансформирована ДНК, нетрансформированные клетки не могут выжить в этой среде. Клетки-реципиенты млекопитающих, используемые для синтеза вариантов по этому изобретению, могут быть выращены во множестве сред. Для выращивания клеток-реципиентов пригодны коммерчески поставляемая среда, такая как 10 (Сигма), минимальная необходимая среда (, (Сигма), -1640(Сигма) и среда Дульбекко, модифицированная по Иглу, (, Сигма). Кроме того, для выращивания клеток-реципиентов как среда может быть использована любая из сред, описанных(.., 58 44 1979),(. ., 102 255 1980), патентах США 4 767 704, 4 657 866, 4 927762 или 4 560 65590/0343087/00195 возобновленном патенте США 30 985. Любая из этих сред как необходимая может быть добавлена с гормонами и/или другими факторами роста (такими как инсулин, трансферрин или фактор роста эпидермиса), солями (такими как хлористый натрий, кальций, магний и фосфат), физиологическими растворами (такими как ), нуклеотидами (такими как аденозин и тимидин), антибиотиками (такими как гентамицин), индикаторными элементами (определяемыми как неорганические составляющие, обычно присутствующие в окончательных концентрациях в микромолярном диапазоне) и глюкозой или эквивалентным источником энергии. Любые другие необходимые добавки могут быть также включены в соответствующих концентрациях, которые должны быть известны квалифицированным специалистам в этой области. Многие эукариотные белки, обычно секретируемые клетками, содержат последовательность эндогенных сигналов как часть последовательности аминокислот. Эта последовательность помечает белок, выводимый из клетки через эндоплазмический ретикулюм и аппарат Гольджи. Последовательность сигналов обычно размещена на аминокислотной терминали белка и занимает длину примерно от 13-ой до 36-ой аминокислот. Хотя действительная последовательность варьирует среди белков, все известные эукариотные последовательности сигналов содержат, по меньшей мере, один положительно заряженный остаток и высокогидро 16 4094 1 фобный участок из 10-15 аминокислот (обычно богатый аминокислотами лейцином, изолейцином, аланином, валином и фенилаланином), расположенный вблизи центра последовательности сигналов. Последовательность сигналов обычно отклоняется от секретируемой формы белка, так как она расщепляется сигнальной пептидазой, расположенной на эндоплазмическом ретикулуме во время перемещения белка в эндоплазмическом ретикулюме. Белок со своей первоначальной последовательностью сигналов часто рассматривают как пребелок, или незрелую форму белка. Однако, не все секретируемые белки содержат расщепленные последовательности сигналов аминокислотных терминалей. Некоторые белки, такие как яичный альбумин, содержат последовательность сигналов,которая размещена во внутренней области белка. Эта последовательность обычно не расщепляется во время транслокации. Белки, обычно находящиеся в цитоплазме, могут быть помечены для секреции путем связывания последовательности сигналов с белком. Это легко осуществляется путем сшивания ДНК, кодирующей последовательность сигналов, с 5-концом ДНК, кодирующей белок, и последующей секреции этого слившегося белка в соответствующую клетку-реципиент. ДНК, кодирующая последовательность сигналов, может быть получена как фрагмент рестрикции из любого гена, кодирующего белок с последовательностью сигналов. Таким образом последовательности прокариотных, дрожжевых и эукариотных сигналов могут быть здесь использованы в зависимости от типа клетки-реципиента, используемой при применении способа по данному изобретению. ДНК, кодирующая часть последовательности сигналов гена, вырезают при использовании соответствующих рестрикционных эндонуклеаз, а затем сшивают с ДНК, кодирующей секретируемый белок, т.е. ТПА. Выбор последовательности функциональных сигналов требует, чтобы последовательность сигналов распознавалась сигнальной пептидазой клетки-реципиента таким образом, чтобы происходило расщепление этой последовательности сигналов и секреция белка. ДНК и последовательность аминокислот, кодирующая часть последовательности сигналов нескольких эукариотных генов, включает, например, человеческий гормон роста, проинсулин и проальбумин, которые известны (см. , , ,1988, .769) и могут использоваться как сигнальные последовательности в соответствующих эукариотных клетках-реципиентах. Последовательности дрожжевых сигналов, как, например, кислой фосфатазы (., . ., 11 1657 1983), альфа-фактора, щелочной фосфатазы и инвертазы(сахарозы), могут быть использованы для непосредственной секреции дрожжевыми клетокамиреципиентами. Последовательности прокариотных сигналов из генов, кодирующих, например,или(.,68 193 1988), ,или ген бета-лактамазы, а также другие гены, могут быть использованы в качестве мишени для белков, поступающих из прокариотных клеток в среду культивирования. В качестве альтернативного варианта получения интересующего секретируемого белка с последовательностью сигналов является химический синтез ДНК, кодирующей последовательность сигналов. В этом способе химически синтезируют обе цепи олигонуклеотидов, кодирующие выбранную последовательность сигналов, а затем отжигают друг с другом для образования двойной цепи. Затем олигонуклетиды в виде двойной цепи связывают с 5-концом ДНК, кодирующей белок. Структуру, содержащую кодирующую белок ДНК, связанную с ним последовательностью сигналов, затем можно связать в подходящем векторе экспрессии. Этот экспрессирующий вектор трансформируют в соответствующую клетку-реципиент, и экспрессируется и секретируется интересующий белок. Культуры клеток-реципиентов млекопитающих и других клеток-реципиентов, которые не имеют жестких клеточных мембранных перегородок, обычно трансформируют с использованием кальциево-фосфатного метода, как было первоначально изложено(, 52 546 1978) и модифицировано,как изложено в разделах 16.32-16.37., выше. Однако, могут быть также использованы другие методы для интродукции ДНК в клетки, такие как(, . . ., 4 1172 1984, слияние протопластов (, . . . . ., 77 2163 1980), электропорация ( . .,., 1 841 1982) и непосредственная микроинъекция в ядра (, , 22479 1980). Дрожжевые клетки-реципиенты, как правило, трансформируют с использованием полиэтиленгликолевого метода, как изложено(. . . .75 1929 1978). Прокариотные клетки-реципиенты или другие клетки-реципиенты с жесткими клеточными стенками преимущественно трансформируют с использованием метода хлористого кальция, как изложено в разделе 1.82 а 1., выше. Альтернативно для трансформации этих клеток может быть использована электропорация. Вариант ТПА предпочтительно выделяют из среды культивирования, как секретируемый белок, хотя он также может быть выделен из лизатов клетки-реципиента, когда он непосредственно экспрессируется без секреторного сигнала. Когда вариант экспрессируется в рекомбинантную клетку, иную, чем клетка человеческого происхождения, он, следовательно, полностью свободен от белка человеческого происхождения. Однако, необходимо очистить вариант от белков рекомбинантных клеток для того, ччтобы получить препа 17 4094 1 раты, которые в значительной степени гомогенны по отношению к белку. На первом этапе среда культивирования или лизат центрифугируют для удаления обломков клеток. Затем вариант очищают от белков с растворенными примесями, например, фракционированием на иммуноспецифичных или ионнобменных колоннах осаждением этанола обратной фазойхроматографией на кремнеземе или на катионообменной смоле, такой какхроматофокусированием - (электрофорезом в полиакриламидном геле - додецилсульфате натрия) осаждением сернокислого аммония или электрофорезом в геле. Также может быть использован ингибитор протеазы, который не препятствует активности ТПА, такой как фтористый фенилметилсульфонил , чтобы препятствовать протеолитической деградации во время очистки, и антибиотики могут быть включены для предотвращения роста добавочных загрязнений. Специалисту, квалифицированному в данной области, понятно, что методы очистки,пригодные для нативного ТПА, могут требовать модификации с учетом изменений характера ТПА или его вариантов при экспрессии в культуру рекомбинантных клеток. В предпочтительной форме получения вариант ТПА выделяют, собранную жидкость культуры клеток фильтруют, и вариант ТПА очищают с использованием лизиноспецифичной хроматографии. Альтернативно,плавающую на поверхности культуру клеток можно пропускать над колонной с предварительным условием наличияи стеклянными шариками, связанными поликлональным антителом А 6 козы с антителами против ТПА, затем колонны уравновешивают буфером и элюцией варианта ТПА. Соединения по настоящему изобретению могут быть получены в соответствии с известными способами приготовления формацевтически полезных составов, в которых продукт ТПА сочетается в примеси с фармацевтически приемлемым носителем. Подходящие носители и их составы описаны в, 16 ., 1980,., изданном. Эти составы обычно содержат эффективное количество варианта ТПА, например, порядка примерно от 0,5 до 5 мг/мл, вместе с соответствующим количеством носителя для приготовления фармацевтически приемлемых составов, пригодных для эффективного назначения пациенту. Вариант ТПА мож начально назначаться пациентам, страдающим сердечнососудистым заболеванием или состоянием, или применять другие способы, которые обеспечивают его доставку в систему кровообращения в эффективной форме. Составы, особенно подходящие для клинического приема вариантов ТПА, используемых при применении настоящего изобретения, включают стерильные водные растворы или стерильные, растворимые в воде порошки, такие как лиофилизированный протеин. Обычно подходящее количество фармацевтически приемлемой соли в составе также используется для придания изотонических свойств. Буфер, например, аргининовое основание, в сочетании с фосфорной кислотой также обычно включается в подходящей концентрации для поддержания соответствующего рН, обычно от 5,5 до 7,5. Дополнительно или альтернативно такая составляющая, как глицерин, может быть включена в состав, чтобы сохранить срок годности при хранении. Дозы и желательные концентрации фармацевтических составов по настоящему изобретению могут изменяться в зависимости от конкретного варианта и конкретного применения. Определение приемлемой дозы находится в компетенции врача, особенно принимая во внимание обширный опыт по назначению рекомбинантного человеческого ТПА немутантного типа. Благодаря своим пролонгированным периодам полураспада плазмы, варианты ТПА по настоящему изобретению особенно пригодны для приема в виде болюса. В соответствии с предпочтительным способом приема, варианты ТПА по настоящему изобретению принимают в начальной внутривенной дозе по одному болюсу при довольно насыщенном механизме очистки ТПА, при необходимости сопровождаемом дальнейшим введением болюса и/или постоянным внутривенным введением. Для вариантов с биологической активностью, сравнимой с активностью ТПА немутантного типа, общая доза составляет преимущественно около 100 мг. Более активные варианты достигают того же эффекта при меньших дозах. Варианты с улучшенной фибриноспецифичностью могут вводиться в больших дозах без существенного увеличения риска осложнения кровотечения. Как правило, дозировка и скорость дозировки могут быть равными или выше, чем обычно при использовании в клинических исследованиях других сердечнососудистых, тромболитических агентов, например,приблизительно 1-2 мг/кг веса тела в качестве внутривенной или внутриартериальной дозы в течение от 1,5 до 12 часов для пациентов (людей), страдающих инфарктом миокарда, легочной эмболией и т.д. Например, при лечении глубоких венозных тромбозов или заболеваний периферических сосудов дозы болюсов порядка 0,05-0,2 мг/кг будут обычно предпочтительны с последующими приемами порядка 0,10,2 мг/кг, вводимыми для поддержания довольно постоянного уровня в крови, предпочтительно порядка 3 мкг/мл. В качестве примера соответствующей формы дозировки может служить ампула, содержащая 50 мг ТПА,аргинина, фосфорной кислоты и полисорбата 80, перемешанных с 50 мл стерильной воды для инъекций и смешанных с соответствующим объемом 0,9 -ной инъекции хлористого кальция. Варианты ТПА по настоящему изобретению также используются для предотвращения отложения фибрина или преобразования адгезии (слипания). Вариант этого использования описан в ЕРО 207 860, опубликованном 4 января 1989 г. В общем, такого рода лечение включает местное (имеющее лишь местное значение) введение состава в сайт потенциального образования фибрина или адгезии, в котором состав содержит тера 18 4094 1 певтически эффективное количество варианта ТПА в малорастворимой форме, который постепенно выделяется в этом сайте в течение примерно от трех дней до двух недель. Обычно вариант ТПА вводят в дозах,достаточных для предотвращения отложения фибрина или образования адгезии, сопровождающих хириргические операции, инфекции, травмы или воспаления. Обычно это количество составляет от 0,02 мг/г геля до 25 мг/г геля, предпочтительно от 0,20 мг/г геля до 2,5 мг/г геля, наиболее предпочтительно примерно от 0,25 мг/г геля до 1,0 мг/г геля. Каждый вариант ТПА, используемый для предотвращения образования адгезии и/или отложения фибрина, обычно включает полутвердый клейкий фармацевтически инертный носитель для размещения фермента в сайте потенциального образования адгезии. Носитель включает длинную цепочку углеводородов или растительных масел и восков, состоящих из смеси модифицированных глицеридов насыщенных и ненасыщенных жирных кислот. Примеры включают полутвердые связующие вещества, такие как вазелин или полусинтетические глицериды, растворители с многими гидроксильными группами, такие как глицерол, длинные цепочки углеводородов, биоразрушаемые полимеры или липосомы. Следующие примеры только иллюстрируют наилучший способ применения согласно изобретению, но никоим образом не ограничивают его объем. ПРИМЕР 1 Получение рекомбинантных вариантов ТПА 1. Конструкция вектора и мутагенез Новые варианты ТПА по настоящему изобретению были сконструированы в векторе экспрессии 7 ТПА (см., например,90/02798, опубликованную 22 марта 1990 г., и 92/02612, опубликованную 20 февраля 1992 г.). Этот вектор, содержащий ТПА мутантного или немутантного типа, использовался для трансфекции и экспрессии в клетках почки человеческого эмбриона (293 С). Направленный на сайт мутагенез ТПА кДНК был выполнен методом., . . ., 13 8765 (1985) с использованием набора, приобретенного у(1253). Для получения желательных мутантов олигонуклеотиды последовательности, кодирующей замещения желательных аминокислот, были синтезированы и использованы в качестве затравок. Эти олигонуклеотиды были отожжены до одиночных рРК 7-ТПА, которые были приготовлены стандартными способами (.,. ., 143 3 (1987). Некоторые из вариантов ТПА были получены с использованием мутагенеза Кункеля ., Т.Д., .. . .82, 488-492 (1985) и , ., . . 154, 367-382 (1987). Смесь трех дезоксирибонуклеотидов дезоксирибоаденозина , дезоксирибогуанозинаи дезоксириботимидинакомбинировали с модифицированным тио-дезоксирибоцитозином, называемым, поставляемым в наборе изготовителем и добавляемым к одноцепочному 7-ТПА, до которого был отожжен олигонуклеотид. При добавлении к этой смеси ДНК-полимеразы была получена цепь ДНК, идентичная рРК 7-ТПА, за исключением мутированных оснований. Кроме того, эта новая цепь ДНК включалавместо ,которая служит для ее защиты от рестрикционного сжигания эндонуклеазами. После того, как матричная цепь двухцепочного гетеродуплекса отмечали соответствующим рестрикционным ферментом, матричную цепь сжигали с нуклеазой Ехо 111 за пределами области, содержащей мутагенный олигомер. Затем реакцию прекращали, чтобы оставить молекулу только частично с одной цепью. Гомодуплексную молекулу ДНК с полной двойной цепью затем получали при использовании полимеразы ДНК в присутствии всех 4 дезоксирибонуклеотидных трифосфатов, АТФ и ДНК-лигазы. Олигонуклеотиды, используемые в качестве затравок для получения молекул рРК 7-ТПА показаны в нижеследующей таблице 1 Таблица 1 Мутация (с одним локусом) 103 105, 107 117(296-299) АААА а) Мутанты с одним локусом - это термин, обозначающий варианты ТПА, которые были получены мутагенезом с одним олигонуклеотидом с использованием в качестве матрицы ТПА немутантного типа. Хотя варианты 105, 107, ТПА и(296-299)АААА ТПА являются многократными мутантами,так как они содержат более одного замещения аминокислот, каждый из них был получен с использованием только одного олигонуклеотида. Это было возможно, поскольку замещенные аминокислоты размещены очень близко друг к другу в полипептидной цепи. 4094 1 Ослабленный вариант вышеописанного способа был использован для приготовления дополнительных многократных мутантов. Для мутантов, описанных ниже, матричный ТПА был не дикого (немутантного) типа (7). Вместо этого используемые матрицы содержали, по крайней мере, одну мутацию, т.е. ДНК, полученную в структуре мутантов в вышеприведенной таблице 1. В таблице 2 ниже указаны ДНК, используемая как матрица, и олигонуклеотид, используемый для получения дополнительных мутаций для каждого полученного мутанта с двойным локусом. Звездочками отмечены варианты, которые иллюстрируют настоящее изобретение. Мутанты (с двойным локусом) 103, 117 105, 107, 117 103, (296-299) 105, 107 ,(296-299) 117, (296-299) б) В качестве матрицы для мутагенеза использовали ДНК, кодирующую мутант ТПА с одним локусом,получив мутантов с двойным локусом. Следующие многократные (с тройным локусом) мутанты были получены, главным образом, следуя предыдущим способам, с использованием многкратных мутантов, указанных в качестве матриц в колонке Матрица. Последовательность каждого олигонуклеотида установлена впереди и сзади в таблице 1 выше. Звездочками отмечены варианты, которые иллюстрируют настоящее изобретение. Мутанты (с тройным локусом) 103, 117, (296-299) 105, 107, 117, (296-299) в) В качестве матрицы для мутагенеза использовали ДНК, кодирующую мутант ТПА с двойным локусом,получив мутантов с тройным локусом. 2. Бактериальная трансформация и приготовление ДНК Структуры мутантов ТПА, получаемые с использованием вышеуказанного протокола, были преобразованы в штамм-реципиент . 294 с использованием стандартной методики( 1.76-1.84., выше) для приготовления и трансформации компетентных клеток. Штамм . 294(который устойчив к фагу Т 1) готовили путем вставки и последующей неточной эксцизии транспозона Т 10 в генеА. Затем этот ген был вставлен с использованием мутагенеза со вставкой транспозона., . ., 116125-159 (1977) в реципиент . ММ 294 (АТТС 31, 446). ДНК экстрагировали из отдельных колоний бактериальных трансформантов с использованием стандартного метода минипрепаратов., выше. Позже плазмиды очищали пропусканием через вращающуюся колонну 6, а затем анализировали секвенированием и сжиганием с помощью рестрикционной эндонуклеазы и электрофорезом в агарозном геле. Альтернативно, последовательность определяли на одноцепочном уровне, и двухцепочные плазмиды очищали от бактериальных трансформантов с помощью набора -плазмид по инструкциям изготовителя. 3. Трансформация клеток линии 293 почек человеческого эмбриона Клетки линии 293 почек человеческого эмбриона выращивали до 70 -ного слияния на 6-луночных планшетах. 2,5 мкг плазмиды, кодирующей мутант ТПА, растворяли в 150 мкл 1 мМ - 0,1 мМ-, 0,227 М . К этому раствору добавляли (капельным методом при интенсивном перемешивании) 150 мкл 50 мМ буферного раствора( 7,35), 280 мМ , 1,5 мМ аРО 4, и при этом допускали образование осадка в течение 10 минут при 25 С. Суспендированный осадок затем добавляли к клеткам в отдельных лунках в 6-луночном планшете и оставляли на всю ночь в термостате. Затем удаляли среду и добавляли 1 мл 20-ного глицерина в(фосфатную соль с физиологическим раствором). Затем добавляли 3 мл свежей среды без сыворотки, и клетки инкубировали в течение 5 дней. Затем среду собирали и анализировали. По настоящему изобретению трансфекция для получения в большом масштабе и очистка вариантов ТПА в клетках 293 линии могут быть выполнены, например, как описано в 90/02798, выше. 4. Экспрессия и очистка в клетках яичника китайского хомяка (СНО) Широко применяемый родительский вектор 65 (трансформированный штамм . АТСС 68 151) использовали для трансфекции и экспрессии вариантов ТПА по настоящему изобретению в клетках яичника китайского хомяка (СНО). Структура 65 описана в патенте США 5 087 572, выданном 11.02.1992 г. 20 4094 1 Клетки - (такие, как описаны, выше), выращенные на неселективнойй среде,трансфектировали вместе с вектором экспрессии на основе 65, в котором последовательность кДНК,кодирующая желательный вариант ТПА, была вставлена в сайт полилинкера, и с вектором 11 с выбором, . . . .80, 2495-2499 (1983) с использованием общей методики, выше. После трансфекции клетки помещали в селективную среду или а) с недостатком глицина, гипоксантина и тимидина, или б) с недостатком этих компонентов и с добавлением метотрексата в концентрациях от 10 до 300 нМ. Для каждой трансфекции изолировали около 50 колоний,предпочтительно с этих планшетов, которые обнаруживали самую высокую точность селекции (с метотрексатом). Клоны размещали в 6-луночных планшетах для анализа экспрессии варианта рекомбинантного ТПА. На сливающиеся лунки действовали средой, свободной от сыворотки (модифицированной из /12 11,содержащей инсулин и трансферрин), в течение 5-6 дней, и ТПА в жидкости клеточной культуры количественно определяли с использованием поликлональной пробы на основе . Для повышения продукции распространяли наилучший продуцирующий клон для каждой трансфекции. Клетки были адаптированы для выращивания в суспензии, и варианты ТПА получили в культурах шелковичного червя с использованием среды без сыворотки. Собранную жидкость клеточной культуры профильтровывали, а затем вариант ТПА был очищали с использованием лизиноспецифичной хроматографии. Пример 2. Характеристика вариантов ТПА 1. Количественное определение ТПА Концентрации белка определяли обычным способом при помощи , стандартизованным для ТПА с нативной последовательностью (см. опубликованный патент ЕРО 93 619). Чистоту и гомогенность белка анализировали путем электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия(-) с буферной системой , , 227 680 (1970). Обычно применяли гели с градиентом от 10 до 20 , и белки визуализировали или с помощью голубого , или с помощью метода протравливания серебром , . . 117307 (1981). Было обнаружено, что при использовании этого метода, приготовленный, как описано выше, вариант ТПА, был чистым и гомогенным. 2. Маркирование вариантов ТПА для изучения очистки и связывания Средство для маркирования активного сайта готовили при помощи радиоиодинации тирозил-пролиларгинил хлорометилкетона , катализированного хлорамином-Т, с использованием модификации метода, описанного 194,495-496 (1962). При обычной реакции в закрытый реакционный сосуд добавляли 50 мкл 1 М - с рН 7,5 к 40 мкл 125 (41,8 нмоль). В этот сосуд добавляли 8,3 мкл(1,8 нмоль в 12 мМ РС). Иодинацию инициировали добавлением 12,5 мкл хлорамина-Т (1 мг/мл в 0,1 М , рН 7,5). Иодинацию подавляли через 60 с при 24 С с помощью 25 мкл метабисульфита натрия (мг/мл в 0,1 М , рН 7,5). Затем реакцию ослабляли путем добавления 2 мл ,и 20 мкл аликвотной пробы разбавленного маркированного реагента добавляли к 1 мл аликвотной пробы культуры супернатанта быстро трансфектируемых клеток линии 293. Эту смесь инкубировали в течение 1 часа, и свободный 125 отделяли от связанного с белком 125 путем фильтрации геля на колонне -10 . Связанный с белком 125, определяли путем осаждения с 10 -ной трихлоруксусной кислоты. Эти условия маркировки оптимизировали для молярного соотношения 111 (йодТПА). Специфическая 125 радиоактивность вариантов ТПА, маркированных 125, составляла около 1 мкС 125 /мкг ТПА. 3. Оценка скорости очистки Очистка плазмы мышей Скорость очистки вариантов ТПА оценивали при помощи радиомаркировки белка, выделяемого клетками линии 293 почек человеческого эмбриона, который через некоторое время инъецировали мышам. В каждом эксперименте использовали по 4 мыши. Мышей разделяли на две группы, и у двух мышей в группе брали кровь в заранее определенные моменты. Данные двух групп были объединены и отмечены на графике зависимости циклов/мин от времени в крови. Участок под кривой расположения плазмыдля каждой мыши определяли с помощью компьютера с использованием последовательных трапеций от 1 до 40 минут. В следующей таблицеобозначает номер эксперимента, используемого для определения скорости очистки.. . представляет скорость очистки, выраженную в мл/мин/кг (веса тела), вычисленную с помощью участка под кривой расположения плазмы, как описано выше. - это коэффициент вариации значения в процентах.. - нормализованный фактор, который представляет собой скорость очистки для варианта ТПА по сравнению с очисткой ТПА немутантного типа (полученного из клеток линии 293). Звездочками отмечены варианты, которые иллюстрируют это изобретение. 4094 1 Таблица 4 Очистка плазмы у мышей Варианты ТПА ТПА немутантного типа 103 105, 107 117 Очистка плазмы у кроликов Фармакокинетический анализ ТПА и вариантов ТПА у кроликов осуществляли с использованием экспериментального протокола модели тромболиза артериально-венозного анастомоза , описанного в разделе 5 ниже. Тефлоновый катетер массой 20 с переключателем потокапомещали в артерию среднего уха анастезированных самцов новозеландских белых кроликов весом 2,5-3,5 кг каждый. Катетер массой 22 с инъекционным наконечником помещали в маргинальную вену противоположного уха. Катетеры промывали и блокировали гепаринизиновым физиологическим раствором. ТПАили варианты ТПА помещали в виде болюса или вливали через венозный катетер. Внутривенное вливание начиналось с 15 -ной дозы болюса, за которой следовало вливание оставшейся 85 -ной дозы в течение 120 минут. Различные дозы ТПАили вариантов ТПА тестировали попротоколу 60 мкг/кг,180 мкг/кг и 540 мкг/кг. Вариант 103,(296-299)АААА тестировали в 4 дозах 20 мкг/кг, 60 мкг/кг, 180 мкг/кг и 540 мкг/кг. Пробы крови брали через 30 минут, 60 минут и 90 минут в течение протокола вливания. Пробы брали через 2, 10, 20, 30, 45, 60, 90, и 120 минут после приема инъекции болюсов. Антикоагулированную плазму была готовили из проб крови, и концентрацию ТПА или варианта ТПА в плазме определяли при помощи поликлонального . Значение скорости очистки для 5 животных оценивали по каждой из многократных доз. Очистка плазмы вариантов ТПА по настоящему изобретению по сравнению с ТПАи известными вариантами ТПА представлена в следующей таблице 5. Звездочкой отмечен вариант, который иллюстрирует это изобретение. Таблица 5 Очистка плазмы у кроликов (мл/мин/кг) Образец ТПА а) Вливание б) Болюсы ТПА 20,73,9 23,96,4(296-299) 20,03,0 н.о. 103 2,70.4 н.о. 103, (296-299) 3,30,2 н.о. 117, (296-299) 22,71,2 н.о. 103, 117, (296-299) 2,10,4 н.о. н.о. - не определяется а) Для тех вариантов ТПА, в которых недостаточные мутации связаны с пониженной очисткой т.е. для немутантного типа и (296-299), очистку плазмы оценивали после длительного внутривенного вливания белка. Концентрацию плазмы в устойчивом состоянии для варианта ТПА определяли в конце вливания с помощью поликлонального . Очистку вычисляли по следующему соотношению Очисткаскорость вливания/концентрация плазмы в устойчивом состоянии. Средние скорости очистки( ) определились повторными анализами (33 и 19) дляи (296-299) соответственно. б) Варианты ТПА с низкими характеристиками очистки (и ТПА ) оценивали у кроликов после приема болюса белка путем внутривенной инъекции. Концентрацию плазмы ТПА в разное время после инъ 22 4094 1 екции определяли с использованием поликлонального . Скорость очистки рассчитывали по соотношению Скорость очисткиДоза/участок под кривой расположения плазмы Средние скорости очистки (. .) определяли повторными анализами (5, 15, 16, 15 и 10) для, 103, 103, (269-299) 117, (296-299) и 103, 117,(296-299) соотвественно. 4.Связывание фибрина Очищенные варианты ТПА из клеток СНО радиомаркированы 125 -и преобразовали в двухцепочную форму с растворимым плазмином при использовании следующей методики. Аликвотные пробы (1,5 мл), содержащие 0,15 мк ТПА, меченого 125 -(или вариантов), инкубировали в фосфатной буферной соли, содержащей альбумин 0,5 бычьей сыворотки и 0,0180, с человеческим плазмином(0,09 единиц казеина в 15 мкл фосфатной буферной соли) в течение 2 часов при 25 С при осторожном встряхивании. Остаточный плазмин был ингибировали апротинином при добавлении 0,9 единиц Т 1 в 15 мкл фосфатной буферной соли. Связывание фибрина оценивали с использованием модификации метода, описанного., 257 2920-2925 (1982). В фосфатной буферной соли готовили растворы лизина и сефарозы, обработанные человеческим фибриногеном, в диапазоне от 0,12 мкг/мл до 8 мг/мл. Аликвоты фибриногена (50 мкл) и ТПА, меченого 125 - , или вариантов ТПА (100 мкл) объединяли в 1,2 -мл полипропиленовых пробирках и быстро перемешивали. Фибриновые тромбы образовывались после добавления человеческого тромбина (50 мкл из 1 един./мл) и наблюдались в течение 60 секунд. После 60 минут при комнатной температуре тромбы центрифугировали при скорости 13000 об/мин в течение 5 минут при 4 С. Аликвоты супернатанта (50 мкл) переносили в отдельные пробирки для подсчета. Чтобы подсчитать общую радиактивность,также учитывали первоначальные полипропиленовые пробирки (содержащие тромбы и оставшуюся жидкость). Количество связанного ТПА было разницей между общей радиактивностью (суммой 50 мкл аликвотной пробы и трубок, содержащих тромбы) и несвязанной радиоактивностью, которая была подсчитана как 4 порядка количества в 50 мкл супернатанта. Результаты проб связывания фибрина, выполненные на обработанном плазмином, очищенном и полученном из СНО материале показаны на фиг. 1 и 2. Цифровые данные связывания фибрина для двухцепочного ТПА (и вариантов ТПА) представлены ниже, в таблице 6. Звездочками отмечены варианты, иллюстрирующие это изобретение. Таблица 6 Пробы связывания фибрина а) Образец ТПА ТПА а) Образцы ТПА, маркированные , обрабатывали плазмином. Эти образцы связывания измеряют фибриноспецифичность ТПА в двухцепочной форме.) Фибрин 50 представляет собой концентрацию фибрина, требуемую для наблюдения 50 образцов белка, связанного с фибрином. с)представляет собой константу кажущегося свойства ТПА связывания с фибриновым тромбом,как определяется по микромолярной концентрации фибрина, требуемого для связывания 50 ТПА. Молекулярный вес фибринового мономера, используемого в этом расчете, был равен 340 килодальтон. Вышеприведенные данные и фиг. 1 свидетельствуют о том, что мутация 103, добавляющая сайт (повышенного) гликозилирования в позиции 103 молекулы человеческого ТПА немутантного типа, вызывает значительные потери связывания с фибрином. Даже если 103 ТПА, как было показано, проявляет примерно в 3-5 раз пониженную очистку по сравнению с человеческим ТПА немутантного типа, его терапевтическое значение незначительно улучшается изза потерь связывания с фибрином. Из предыдущих данных, представленных на фиг. 2, видно, что связывание фибрина 103 ТПА улучшается главным образом при осуществлении второй мутации, вызывающей отсутствие гликозилирования в по 23 4094 1 зиции 117 молекулы человеческого ТПА немутантного типа. Более того, вариант ТПА 103, 117 сохраняет свою пониженную очистку по сравнению с человеческим ТПА немутантного типа (или ТПА ). Осуществление дальнейшей мутации в позициях 296-299 молекулы человеческого ТПА немутантного типа, который, как известно, дает значительное улучшение фибриноспецифичности (296-299),генерирует тройной мутант, который в значительной степени сравним с человеческим ТПА немутантного типа, благодаря его существенно пониженной скорости очистки и повышенной фибриноспецифичности без значительных потерь связывания с фибрином. 5. Проба лизиса тромба(Модель тромболиза артерио-венозного анастомоза у кролика) Тромбы получали - путем помещения 0,2 мл или свежей 70 -ной цельной крови кролика (разбавленной 0,15 М ), или плазмы, богатой тромбоцитами, собранной с помощью(этилендиаминтетрауксусной кислоты) у кроликов (0,8 х 106 тромбоцитов/мкл) в цилиндр 0,5 мл шприца. К плазме, богатой тромбоцитами, добавляли человеческий тромбин и 2 (конечные концентрации 0,2 мкг/мл и 15 мМ соответственно). Аликвотные пробы цельной крови и плазмы, богатой тромбоцитами, перед переносом в шприц нейтрализовали человеческим фибриногеном с 125. Длинную хлопчатобумажную нитку пропускали через цилиндр шприца. Эта нить служила для осаждения образующихся тромбов. После того, как тромбы инкубировали при 37 С в течение одного часа, поршень вынимали и в шприцы (1 с цельной кровью и 1 с плазмой) вставляли трубки из силастика. Эти трубки присоединяли к катетерам, которые предварительно имплантировали в сонную артерию и яремную вену новозеландского белого кролика весом 2,5-3,0 кг. Кровь пропускали путем артериальной циркуляции через тромб и возвращали по венозному руслу. Кровяной поток через анастомоз составлял примерно 20 мл/мин. ТПАвводили через венозный катетер путем вливания в течение 90 минут (15 доза загрузки). 103, 117, (296-299) вводили как внутривенный болюс. Также вводили гепарин (300 ед/кг) в виде болюса тем же путем при -10 и 45 минутах. Тромболиз контролировали внешним гамма-детектором через 120 минут времени проведения эксперимента. Определяли относительную активность, исходя из полулогарифмического графика характеристики дозы для мутантов относительно стандартной кривой ТПА . По окончании экспериментов вынимали цепь тромбов, и катетеры заполняли физиологическим раствором и закупоривали гепарином (500 ед./катетер). Десять животных использовали один раз в день в течение 5 последовательных дней. Результаты приведены на фиг. 3-6. Данныепоказывают, что лизис тромбов осуществляет тройной мутант 103, 117, (296299) ТПА значительно быстрее, чем ТПА , как в тромбах из цельной крови, так и в тромбах из плазмы, богатой тромбоцитами (фиг. 3 и 4). Дальнейшие данные показывают, чтоактивность 103, 117, (296-299) ТПА на 6,0 порядков и 9,5 порядков выше, чем ТПАв тромбах из цельной крови и тромбах из плазмы, богатой тромбоцитами, соответственно (фиг. 5 и 6). Эти числовые данные определяли на основе кривых параллельных линий, соответствующих экспериментальным точкам, нанесенным на фиг. 5 и 6. Депонирование материалов Клетки . 294, трансформированные вектором 5, депонировали АТСС (Американской коллекцией видов культур), 12301, , ,(АТСС) 25 октября 1989 г. и были внесены в каталог АТСС под 68 151. Депонирование осуществляли на основе положений Будапештского договора о международном признании депонирования микроорганизмов в целях процедуры патентования и нижеследующих правил (Будапештского договора). Оно обеспечивает хранение жизнеспособной культуры в течение 30 лет с даты депонирования. Организм будет в распоряжении АТСС по условиям Будапештского договора и подчиняться соглашению между , . и АТСС, который обеспечивает постоянную и неограниченную доступность потомков культуры для общественности по выдаче соответствующего патента США или по выкладке любой заявки США или иностранной, какая придет первой, и обеспечивает доступность потомков лицам, указанным Комиссаром США по патентам и товарным знакам, которым дано на это право согласно статье 35(свода законов США), параграф 122 и правилам Комиссара, относящимся к этому(включая 37 параграф 1.14 с конкретной ссылкой на 886638). Все ограничения на распространение АТСС 68151 были сняты 11 февраля 1992 г. Представитель по настоящей заявке согласился, что если культура на депонировании умрет или будет потеряна, или разрушена, то, выращенная при подходящих условиях, она будет срочно заменена по извещении жизнеспособным образцом той же культуры. Доступность депонированного штамма не должна использоваться как лицензия на применение изобретения, в нарушение прав, представленных представителями правительства, в соответствии с его патентными законами. Предшествующее письменное описание считается достаточным, чтобы квалифицированный специалист в данной области мог осуществить изобретение. Настоящее изобретение не ограничивается областью депонирования конструкции. Депонирование материала не допускает ни то, что письменное описание, заключающееся здесь, неадекватно возможности применения любого аспекта изобретения, включая наилучший способ его применения, ни то, что его следует рассматривать, как ограничивающее сферу притязаний до специфического иллюстрирования, которым оно представлено. 24 4094 1 Хотя предшествующее относится к конкретным предпочтительным вариантам выполнения, понятно, что настоящее изобретение не ограничивается этим. Для обычных специалистов в данной области может случиться, что различные модификации могут быть осуществлены для описанных вариантов выполнения без отклонений от общей концепции изобретения. Все такие модификации предназначены находиться в пределах сферы настоящего изобретения. ХАРАКТЕРИСТИКА ПОСЛЕДОВАТЕЛЬНОСТЕЙ Название изобретения Варианты гликозилирования тканевого плазминогенного активатора с улучшенными тераптевтическими свойствами(А) Тип носителя 5,25 дюйм, 360 Кб гибкий диск (В) Компьютер совместимый сС) Управляющая система -/ Программное обеспечение(С) Номер квитанции об уплате пошлины 757 Описание последовательности обозначение последовательности 124 Описание последовательности обозначение последовательности 230 Описание последовательности обозначение последовательности 324 Описание последовательности обозначение последовательности 436 Государственный патентный комитет Республики Беларусь. 220072, г. Минск, проспект Ф. Скорины, 66. 26
МПК / Метки
МПК: C12N 9/64, C12N 5/10, C12N 15/56, A61K 38/48
Метки: плазминогенного, способ, кодирующие, связывания, тпа, тканевого, восстановления, составы, варианты, лечения, активатора, последовательности, фибрина, днк, человеческого, способы, свойства
Код ссылки
<a href="https://by.patents.su/26-4094-varianty-chelovecheskogo-tkanevogo-plazminogennogo-aktivatora-tpa-posledovatelnosti-dnk-kodiruyushhie-varianty-tpa-sostavy-sposoby-lecheniya-i-sposob-vosstanovleniya-svojjstva-svya.html" rel="bookmark" title="База патентов Беларуси">Варианты человеческого тканевого плазминогенного активатора (ТПА), последовательности ДНК, кодирующие варианты ТПА, составы, способы лечения и способ восстановления свойства связывания фибрина с ТПА</a>
Способ активации тканевого активатора плазминогена (его варианты)

Номер патента: 526
Опубликовано: 30.06.1995
Авторы: Ральф Маттес, Штефан Фишер, Райнер Рудольф
МПК: C12N 15/12, C12P 21/00
Метки: плазминогена, активатора, его, активации, тканевого, способ, варианты
Текст:
...(100) в 0,1 молъ/л трис/НС 1(рН 10,5) 1 мысль/л ЕДТА, 0,5 мо.пъ/л Ьаргинииа, 1 мг/мл ВЗА, 0,5 мол/л СЗН (восстановленный глютатион),0,5 моль/л 0530 (дисупъфид глютатиона). . 1. Зависмость выхода активации от добавки Ь-аргинина или гуанидингидрохпорнда. .Зависимость выхода активации от КОНЦВНТРЗЦНН РЗРГННННЗ ПРЕДСТЕВЛЕ нав таби. 2.- 2 . Зависимость активации от добавкиЗависмостъ актиацииот добавки мочевины следующаяМочерина Активность,5...
Производные 5-амино-8-метил-7-пирролидинилхинолин-3-карбоновой кислоты, способы их получения (варианты), фармацевтическая композиция, способ лечения, промежуточный продукт

Номер патента: 3779
Опубликовано: 30.03.2001
Авторы: Синго Ясуда, Тошихико Йошида, Нориюки Кадо, Йоичи Ямамото, Хидео Като, Ясуо Ито
МПК: A61K 31/47, C07D 401/04, A61K 31/40...
Метки: способы, фармацевтическая, варианты, лечения, 5-амино-8-метил-7-пирролидинилхинолин-3-карбоновой, кислоты, производные, композиция, продукт, получения, промежуточный, способ
Текст:
...активность,столь же высокую, что и у аналогичного ссылочного соединения С (имеет только один отличающийся заместитель в 5-положении, чем и отличается от соединения настоящего изобретения), и, кроме того, не показывает токсичности (хромосомная аберрация), наблюдаемой у ссылочного соединения С. Хотя ссылочное соединение С, как и соединение настоящего изобретения, не проявляет фототоксичности, введение ссылочного соединения С вызвало...
Способ получения дорамектина (варианты), производные дорамектина, способ получения производного дорамектина, способы лечения (варианты).

Номер патента: 3760
Опубликовано: 30.12.2000
Авторы: Чарльз Уильям МУРТИАШОУ, Константин СКЛАВОУНОС, Томас Чарльз КРОУФОРД, Стефен П. ГИБСОН, Нейл ДЕМЕРС
МПК: A61P 33/10, A01N 43/90, A61K 31/70...
Метки: дорамектина, способ, производного, варианты, лечения, получения, способы, производные
Текст:
...способами органической химии. 9 3760 1 Соединения общей формулы , согласно данному изобретению, где 2 - Н и 1 - (С 2-С 5)-алканоил или феноксиацетил, легко получают в соответствии со способом данного изобретения. Соединение формулыреагирует с ацилирующим агентом в инертном растворителе в присутствии акцептора протона, такого как пиридин,4-диметиламинопиридин,пиперидин,пирролидин,триэтиламин,морфолин или ди(изопропил)этиламин, но не...
Фрагмент ДНК, кодирующий полипептид К2Р со свойствами активатора плазминогена, полипептид К2Р со свойствами активатора плазминогена, рекомбинантная плазмидная ДНК рА27.3, кодирующая полипептид К2Р со свойствами активатора плазменогена и фармацевтическая

Номер патента: 2120
Опубликовано: 30.06.1998
Авторы: УЛЬРИХ КОНЕРТ, УЛЬРИХ МАРТИН, Райнер Рудольф, ШТЕФЕН ФИШЕР, АННЕ ШТЕРН
МПК: C12N 15/58, A61K 37/47, C07K 14/745...
Метки: полипептид, плазменогена, ра27.3, свойствами, кодирующая, плазминогена, фармацевтическая, плазмидная, днк, активатора, к2р, кодирующий, рекомбинантная, фрагмент
Текст:
...25 С. После установления рН-значения 3 с помощью 1 (25) осуществляют диализ против 10 ммоль/л 1 (3 х 100 л 48 часов, 4 С). После диализа центрифугируют и прозрачную надосадочную жидкость обрабатывают далее. Натурирование. Реакционный сосуд емкостью 10 л заполняют 0,1 ммоль/л Трис-1, 0,8 ммоль/л -аргинина, 2 ммоль/л(гентатион, восстановленная форма), 1 ммоль/л ЭДТК, 8,5. Ренатурирование проводят при 20 С путем трехкратной добавки, смотря по...
Способ прямого восстановления содержащего оксиды железа материала (варианты) и устройство для осуществления способа (варианты)

Номер патента: 3196
Опубликовано: 30.12.1999
Авторы: УИПП, Рой, Хуберт, Мл., ЦИП, Герхард, МИЛИОНИС, Константин, ГУШЕХ, Мортеза Садат
МПК: C21B 13/00, C21B 13/02
Метки: способ, железа, варианты, содержащего, оксиды, материала, способа, восстановления, устройство, прямого, осуществления
Текст:
...прошедшего очистку от СО 2, примешивают к колошниковому газу, подвергнутому очистке от СО 2. Часть колошникового газа, не прошедшего очистку от СО 2, примешивают к колошниковому газу, подвергнутому очистке от СО 2. Колошниковый газ, подвергнутый очистке от СО 2, могут смешивать с одним или несколькими следующими газами в качестве газов синтезаотходящий газ, отходящий газ,доменный газ от доменных печей,колошниковый газ от...
Предыдущий патент: Трансдермальный препарат продленного действия с антиостеопоротической активностью
Следующий патент: Клеящий штифт и способ его получения
Случайный патент: Способ оценки способности человека управлять перемещением центра тяжести своего тела