Способ интрогрессии генов аллотетраплоидного картофеля дикого вида в геном картофеля Solanum tuberosum
Номер патента: 11631
Опубликовано: 28.02.2009
Авторы: Воронкова Елена Васильевна, Ермишин Александр Петрович, Лисовская Вероника Михайловна





Текст
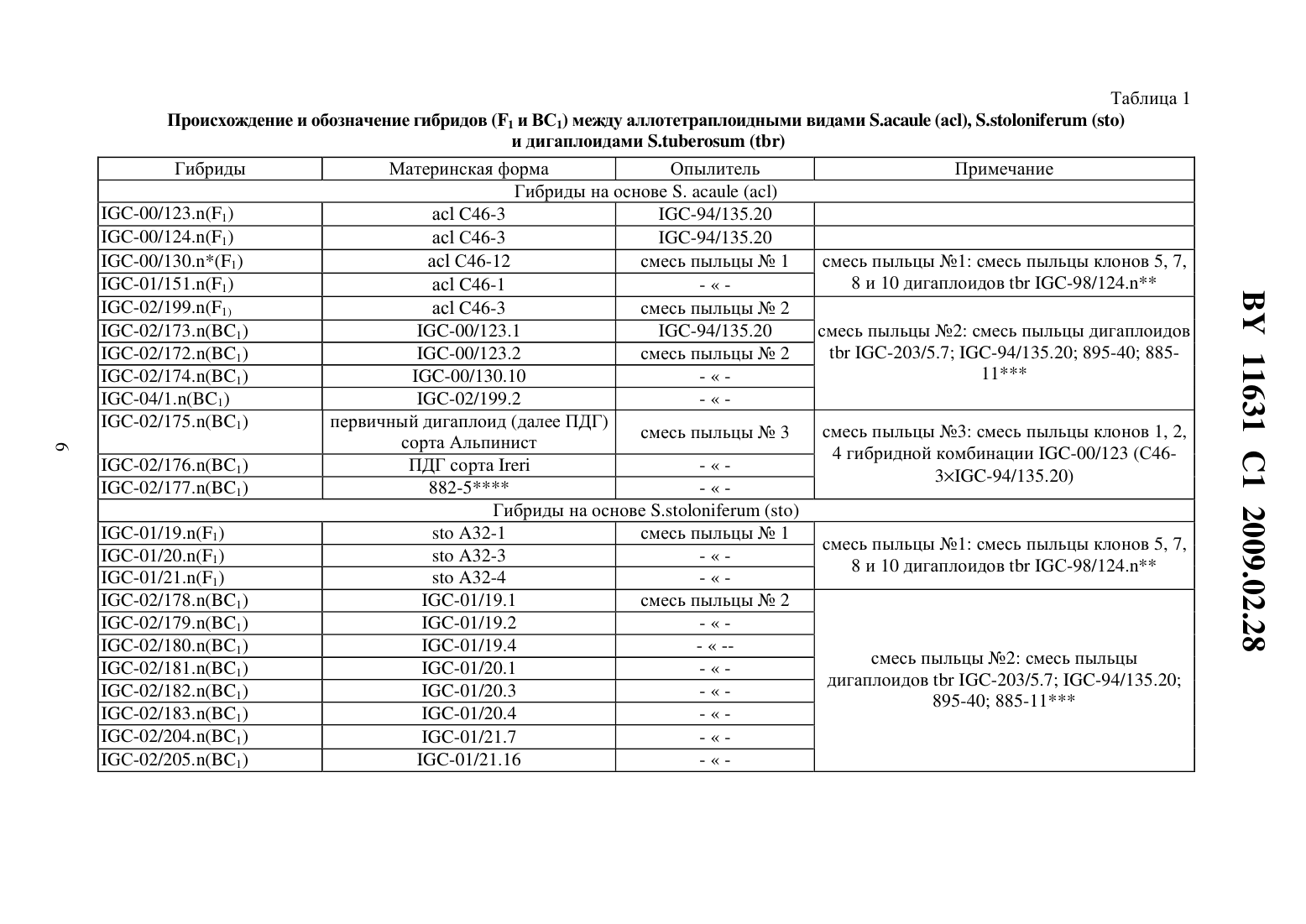
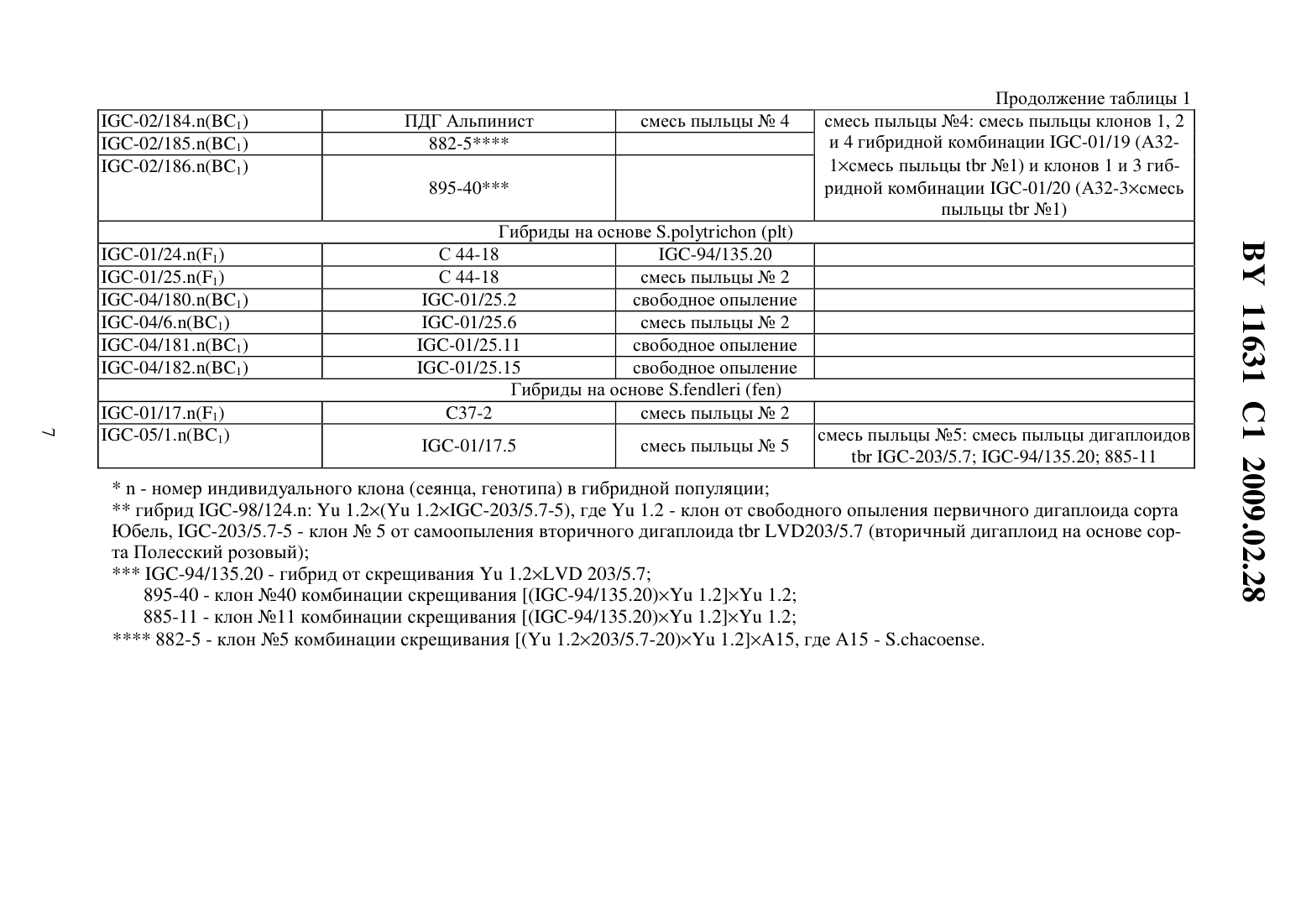
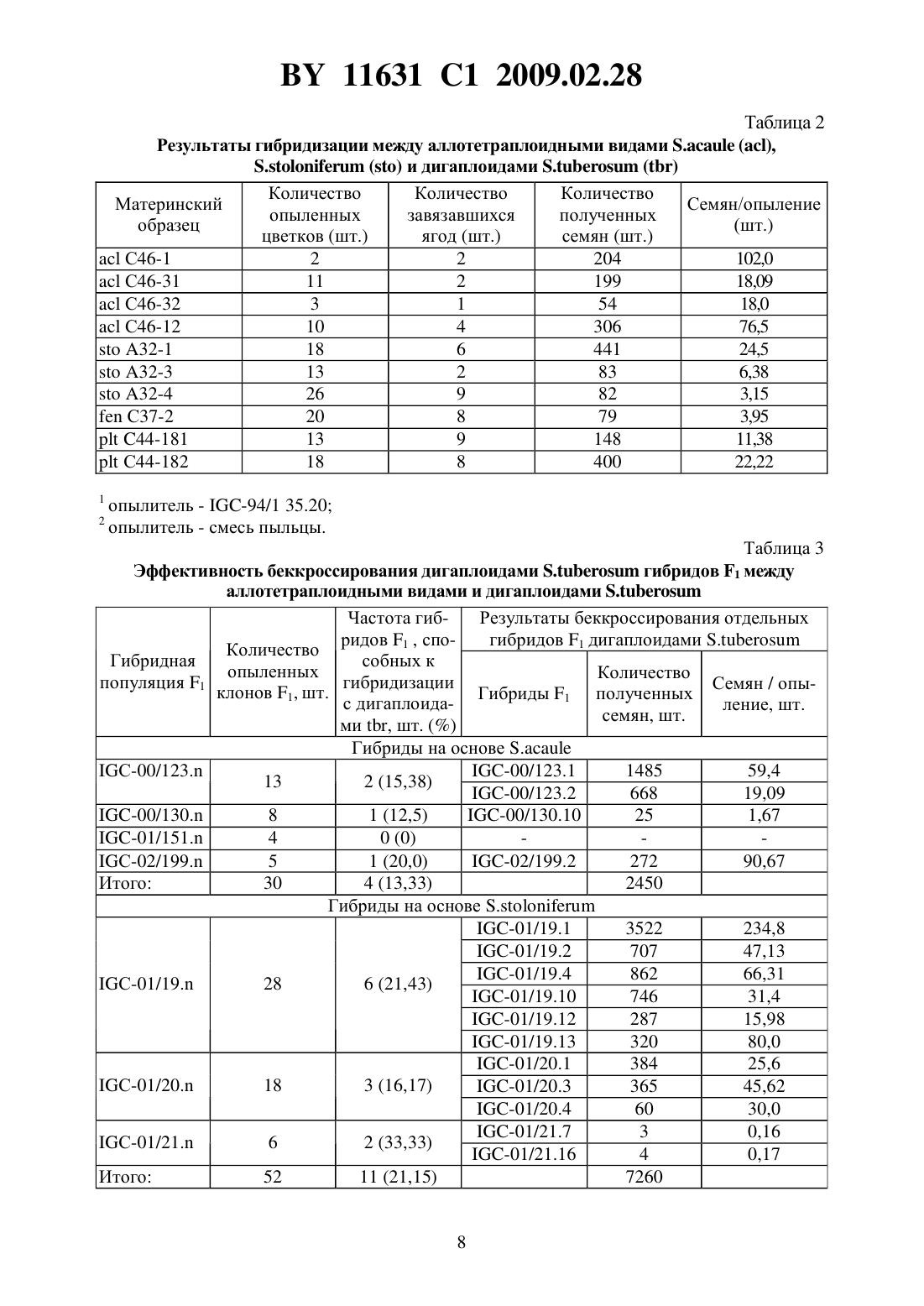
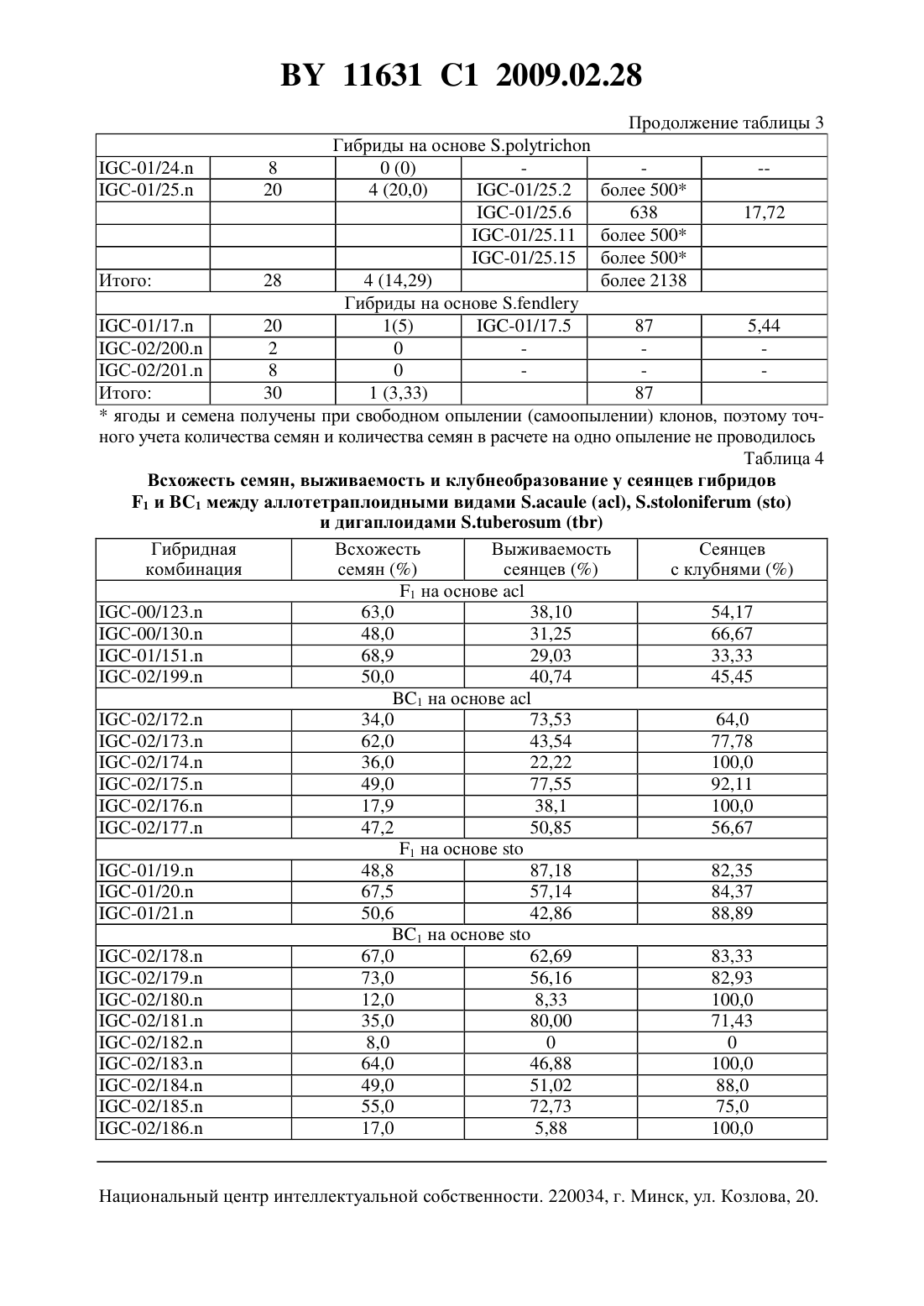
опылению происходит завязывание ягод, содержащих семена, полученные 1) в результате гибридизации между 5 о 1 а 1111111 шЬегозиш и 5 о 1 а 1111111 р 11111 еа (их можно отличить по наличию маркерного признака) 2) семена, полученные в результате развития неоплодотвореннь 1 х яйцеклеток (дигаплоидь 1 5 о 1 а 1111111 шЬегозиш) и 3) семена, полученные в результате гибридизации между 5 о 1 а 1111111 шЬегозиш и 5 о 1 а 1111111 асаи 1 е. Последние являются недоразвитыми из-за межвидовой несовместимости. Для того чтобы их сохранить, применяют метод культуры зародышей 111 что. Недостатками известного способа являются сложность и невысокая результативность метода культуры зародышей 111 уйго. Отбор по селекционноценным признакам (интрогрессия селекционно-ценных генов дикого вида), элиминация неблагоприятных генов дикого вида проводятся среди тетраплоидного селекционного материала, для которого характерно сложное расщепление. В литературе отсутствуют сведения о применении этого способа для получения межвидовых гибридов между культурным картофелем 5 о 1 а 1111111 шЬегозиш и другими (помимо 5 о 1 ап 11111 аса 111 е) аллотетраплоидными дикими видами картофеля.Известен способ получения гибридов между культурным тетраплоидным картофелем 5 о 1 а 1111111 шЬегозиш и аллотетраплоидными дикими видами картофеля, в котором с целью преодоления нескрещиваемости между видами применяют удвоение хромосом у аллотетраплоидного вида, полученные октоплоиды скрещивают с тетраплоидным культурным картофелем, затем полученные гексаплоидные гибриды несколько раз беккроссируют культурным картофелем до достижения тетраплоидного уровня селекционного материала 3. Этот способ имеет следующие недостатки. Во-первых, присутствие у полиплоидных гибридов полного набора хромосом дикого вида затрудняет рекомбинацию генов дикого и культурного видов, в силу того что в мейозе будет иметь место конъюгация, прежде всего, между гомологичными хромосомами одного вида. Во-вторых, требуется достаточно длительное время для снижения уровня плоидности селекционного материала до тетраплоидного уровня. В-третьих, отбор по селекционно-ценным признакам, элиминация неблагоприятных генов дикого вида проводятся среди полиплоидного и анеуплоидного селекционного материала, для которого характерно сложное расщепление.Известен способ получения гибридов между культурным тетраплоидным картофелем 5 о 1 а 1111111 шЬегозиш и аллотетраплоидными дикими видами картофеля, который включает получение соматических гибридов между культурным тетраплоидным картофелем 501 ап 11111 шЬегозиш и дигаплоидами (11 2 х 24) аллотетраплоидных диких видов, например 5 о 1 а 1111111 аса 111 е, многократное беккроссирование полученных гексаплоидных гибридов до достижения тетраплоидного уровня селекционного материала, в ходе которого проводят отбор по признакам, которые предполагают перенести в селекционный материал от дикого вида 4. Этот способ имеет следующие недостатки. Во-первых, методы получения дигаплоидов аллополиплоидных видов, получения соматических гибридов путем слияния протопластов являются сложными, требуют дорогостоящего оборудования и высокой квалификации персонала. Во-вторых, требуется достаточно длительное время для снижения уровня плоидности селекционного материала до тетраплоидного уровня. В-третьих, отбор по селекционно-ценным признакам, элиминация неблагоприятных генов дикого вида проводятся среди полиплоидного и анеуплоидного селекционного материала, для которого характерно сложное расщепление.Наиболее близким техническим решением является способ получения гибридов между культурным картофелем 5 о 1 а 1111111 шЬегозиш и аллотетраплоидными дикими видами картофеля, включающий гибридизацию между аллотетраплоидными видами картофеля и дигаплоидами (211 2 х 24) 5 о 1 а 1111111 шЬегозиш, удвоение хромосом у полученных триплоиднь 1 х гибридов, многократное беккроссирование полученных гексаплоидных гибридов до достижения тетраплоидного уровня селекционного материала, в ходе которого проводят отбор по признакам, которые предполагают перенести в селекционный материал от дикого вида 5. Этот способ имеет следующие недостатки. Требуется достаточно дли ВУ 11631 С 1 2009.02.28тельное время для удвоения числа хромосом триплоидных гибридов, мейотическое удвоение в большинстве случаев оказывается неудачным. Требуется достаточно длительное время для снижения уровня плоидности селекционного материала до тетраплоидного уровня. Отбор по селекционно Ценным признакам, элиминация неблагоприятных генов дикого вида проводятся среди полиплоидного и анеуплоидного селекционного материала,для которого характерно сложное расщепление.Сущность изобретения состоит в решении задачи по упрощению процедуры получения первого беккроссного поколения гибридов между культурным картофелем 5 о 1 аш 1 ш шЬегозиш и аллотетраплоидными дикими видами картофеля и повышению эффективности отбора по селекционно ценным признакам и элиминации неблагоприятных генов дикого вида среди селекционного материала на основе полученных межвидовых гибридов.Решение задачи состоит в том, что в известном способе, включающем гибридизацию между аллотетраплоидными видами картофеля и дигаплоидами 5 о 1 апиш гиЬегозиш, отбирают среди гибридов между аллотетраплоидными видами картофеля и дигаплоидами 501 ап 11 ш шЬегозиш диплоидные генотипы, способные повторно скрещиваться с дигаплоидами 5 о 1 аш 1 ш гиЬегозиш, отбор по признакам, которые предполагают перенести в селекционный материал от дикого вида, и элиминацию неблагоприятных генов дикого вида проводят среди полученного диплоидного селекционного материала.Сопоставительный анализ заявляемого решения с прототипом показывает, что заявляемый способ отличается от известного тем, что исключает применение сложной и длительной процедуры удвоения хромосом полученных межвидовых триплоидных гибридов,сложной и длительной процедуры многократного беккроссирования полученных гексаплоиднь 1 х гибридов до достижения тетраплоидного уровня селекционного материала. С помощью простой процедуры гибридизации между первичными межвидовыми гибридами и дигаплоидами 5 о 1 апиш шЬегозиш несложно выявить диплоидные формы среди межвидовых гибридов (они завязывают с высокой частотой ягоды с жизнеспособными семенами). Полученные диплоидные межвидовые гибриды первого поколения (131) имеют регулярный мейоз, достаточно высокую фертильность, что дает возможность повторно эффективно скрещивать их с дигаплоидами 5 о 1 ап 11 ш гиЬегозиш как в качестве материнских форм, так и опылителей. Диплоидные межвидовые гибриды 131 и последующие поколения беккросса характеризуются высокой стабильностью генома (2 п 2 х 24). Отбор по селекционно ценным генам и элиминация неблагоприятных генов дикого вида производятся среди эуплоидного диплоидного селекционного материала, для которого характерно простое дисомное наследование признаков.Табл. 1 представляет сведения о происхождении и номенклатуре родительских образцов, а также комбинаций межвидовых гибридов (131 и ВС 1), которые были использованы и получены при реализации предлагаемого способа получения гибридов между культурным картофелем 5 о 1 апиш шЬегозиш и аллотетраплоидными дикими видами картофеля.Табл. 2 представляет результаты гибридизации между рядом клонов диких аллотетраплоиднь 1 х видов 5 о 1 аш 1 ш аса 111 е, 5 о 1 апиш 5 го 1 оп 1 Гегиш, 5 о 1 аш 1 ш ро 1 упг 1 с 11 оп, 5 о 1 аш 1 ш Гешпегй и дигаплоидами 5 о 1 аш 1 ш гиЬегозиш, которые свидетельствуют о возможности получения достаточно большого количества выполненных нормально развитых гибридных семян 131.Табл. 3 представляет результаты гибридизации между полученными межвидовыми гибридами 131 и дигаплоидами 5 о 1 аш 1 ш шЬегозиш, которые показывают что образцы, способные завязывать семена при опылении пыльцой диплоидного картофеля, можно обнаружить во многих комбинациях 131 с достаточно высокой частотой (до 33,3 ).Табл. 4 представляет результаты оценки всхожести семян, выживаемости сеянцев и клубнеобразования у межвидовых гибридов 131 и первого поколения беккросса межвидовых гибридов, полученного путем скрещивания 131 с дигаплоидами 5 о 1 аш 1 ш гиЬегозиш (как в качестве материнских форм, так и в качестве опылителей). Эти результаты свидетельст ВУ 11631 С 1 2009.02.28вуют о высокой эффективности элиминации неблагоприятных генов дикого вида в селекционном материале на диплоидном уровне.Предлагаемый способ получения гибридов между Культурным картофелем 5 о 1 апиш шЬегозиш и аллотетраплоидными дикими видами картофеля реализован следующим образом. В качестве исходного материала для гибридизации аллотетраплоидов с диплоидным 5111 Ьего 5 иш были использованы растения картофеля аллотетраплоидных видов 5 о 1 аш 1 ш асаи 1 е (Не образца по каталогу Всероссийского института растениеводства им. Н.И.Вавилова (далее Не по каталогу ВИР) К-4256 - представлен тремя клонами С 46-1, С 46-3 и С 4612), 5.5 о 1 оп 1 Гегиш (Не по каталогу ВИР К-17152 - представлен тремя клонами А 32-1, А 32-3 и А 32-4), 5.ро 1 уг 1 с 11 оп (Не по каталогу ВИР К-20086 - представлен одним клоном С 44-18),5.Геп 11 егу (Не по каталогу ВИР К-5747 - представлен одним клоном С 37-2), предоставленные лабораторией биотехнологии Института картофелеводства НАН Беларуси. Родительские клоны поддерживаются в культуре 111 уйгго путем периодического, один раз в 30 дней,черенкования пробирочных растений. В работе использованы диплоидные формы 5111 Ьего 5 иш с высокой фертильностью пыльцы, полученные в лаборатории генетики картофеля Института генетики и цитологии НАН Беларуси (табл. 1).Для проведения гибридизации растения диких видов высаживают в грунт оранжереи из культуры 111 уйгго в начале мая. Растения высаживают рядами расстояние между рядами 70 см, между растениями в ряду 50 см. Режим выращивания растений температура воздуха 20-25 С - днем, 15-20 С - ночью, влажность 70 , освещение естественное. Для стимуляции цветения растения, достигшие высоты 35-40 см, подвязывают к кольям,обнажают от почвы место отхождения корней от стебля. Образующиеся столоны обрь 1 вают или оставляют над поверхностью почвы. Аллотетраплоиды используют в качестве материнских форм. Накануне опыления проводят кастрацию бутонов, начинающих приобретать окраску (нижняя часть бутонов в это время имеет белый цвет, верхняя часть бутонов - цвет, характерный для цветков опыляемого образца), путем удаления пыльников с помощью пинцета. Также удаляют в соцветии все раскрытые цветки, полураскрытые и незрелые (зеленые или начинающие приобретать беловатую окраску) бутоны.Сбор пыльцы дигаплоидов 5.шЬего 5 иш проводят следующим образом распустившиеся цветы собирают, подсушивают 1 сутки при комнатной температуре, затем вытряхивают из них пыльцу с помощью электрического вибратора, например электробритвы типа Микма 100, 101, Эра и др. Собранную пыльцу переносят в лунки пластмассовых планшетов, применяемых для иммуноферментного анализа, или пластиковые одноразовые чашки Петри диаметром 5 см. Опыление кастрированных цветков диких видов осуществляют свежесобранной пыльцой или пыльцой, хранившейся в морозильной камере при температуре -10 С не более 5 дней путем погружения рыльца пестика в пыльцу. На опыленное соцветие вешают этикетку, в которой указана дата опыления и наименование родительских форм. Применяют опыление смесью пыльцы дигаплоидов 5111 Ьего 511 ш или опыление пыльцой одного генотипа дигаплоидов 5. шЬегозиш. Смесь пыльцы готовят непосредственно перед гибридизацией путем объединения и тщательного перемешивания в лунке равных объемов пыльцы разных генотипов дигаплоидов 5111 Ьего 511 ш.Через месяц после опыления на завязавшиеся ягоды одевают марлевые мешочки и подвязывают их к стеблю материнского растения. Гибридные семена извлекают из созревших ягод с помощью скальпеля или пинцета, подсушивают и хранят в бумажных пакетах при комнатной температуре. Для определения всхожести семян их замачивают на сутки в 0,1 растворе гиббереллина и высевают в начале апреля в плошки с торфяной почвой Двина, смоченной водой. Глубина заделки семян в почву 0,5 см. Через двадцать дней после посева учитывают количество взошедших семян. Рассчитывают всхожесть семян как отношение числа взошедших семян к числу посеянных. Выживаемость сеянцев определяют через шесть недель после посева как число выживших сеянцев к числу взошедших семян.Выжившие сеянцы гибридов 131 между аллотетраплоиднЬ 1 ми видами и дигаплоидами 5111 Ьего 5 иш пересаживают в теплицы и выращивают, как описано выше, с целью их гибридизации, как описано выше, с дигаплоидами 5111 Ьего 5 иш. При этом ягоды завязывают только те Межвидовые гибриды, Которые являются диплоидами (2 п 2 х 24). Их доля в популяции гибридов 131 между аллотетраплоиднЬ 1 ми видами и дигаплоидами 5.11 Ьего 5 иш составляет, в зависимости от генотипа материнской формы, от О до 33,3 (табл. 3). Наибольшая частота диплоидных межвидовых гибридов 131 наблюдается у 5.5 о 1 о 111 Гегиш (от 16,2 до 33,3 ), наименьшая - 5.Ге 1111 егу (от О до 5 ).Тем не менее, у всех проанализированных аллотетраплоидных видов можно выделить генотипы, образующие такие генотипы, что позволяет получить достаточное количество семян первого поколения беккросса для последующей селекции на диплоидном уровне с целью интрогрессии ценных генов диких аллотетраплоидных видов в селекционный материал 5111 Ьего 511 ш.Из данных табл. 4 видно, что первое поколение беккросса существенно превосходит 131 по выживаемости сеянцев и клубнеобразованию в условиях умеренных широт, что свидетелЬствует о высокой эффективности элиминации неблагоприятных генов дикого вида в селекционном материале на диплоидном уровне.2. Негшзеп 1.6.Т 11., Уегбепшз 1. 5 е 1 есг 1 оп Егош 5 о 1 а 1111 ш шЬегозиш гоир Р 1111 геа оГ депоурез сошЬ 1 п 1 п 3 111311 Ггечиепсу 11 ар 1 о 11 1 п 111 с 1 оп 91/1111 11 ошо 2 у 3 о 51 у Гог ешЬгуо-зрог. Еир 11 уг 1 са, 1973. У. 22. - Р. 244-259.3. Ьашш К. 111 уе 513 а 1 оп оГ зоше иЬег-Ьеагш 5 о 1 а 1111 ш 11 уЬг 115. Неге 11 а 5, 1953. У. 39. Р. 97-112.
МПК / Метки
МПК: A01H 1/02
Метки: генов, интрогрессии, solanum, способ, вида, дикого, tuberosum, аллотетраплоидного, геном, картофеля
Код ссылки
<a href="https://by.patents.su/9-11631-sposob-introgressii-genov-allotetraploidnogo-kartofelya-dikogo-vida-v-genom-kartofelya-solanum-tuberosum.html" rel="bookmark" title="База патентов Беларуси">Способ интрогрессии генов аллотетраплоидного картофеля дикого вида в геном картофеля Solanum tuberosum</a>
Способ получения фитофтороустойчивого картофеля

Номер патента: 9691
Опубликовано: 30.08.2007
Авторы: Семанюк Тамара Владимировна, Яковлева Галина Анатольевна
МПК: A01H 1/02, C12N 15/02
Метки: фитофтороустойчивого, получения, способ, картофеля
Текст:
...и 0,510,которую предварительно центрифугируют, образовавшиеся визуально обнаруживаемые индивидуальные колонии переносят на поверхность агаризованной среды СТ 1, индукцию морфогенеза проводят при температуре 18-20 С, а для укоренения побегов используют среду Мурасиге-Скуга, содержащую 10 г/л сахарозы, 1 мг/л витамина В 1, 0,5 мг/л витамина В 6, 0,25 мг/л эпибрассенолида, 0,5 мг/л гиббереллина 3, 0,5 мг/л аденина и 0,2 мг/л 3-индолилмасляной...
Способ выращивания картофеля

Номер патента: 54
Опубликовано: 30.09.1994
Автор: Иващенко Алексей Иванович
МПК: A01C 21/00
Метки: способ, картофеля, выращивания
Текст:
...и хлористого калия внос-или вразброс под препосевную культивацию с помощью разр брасывателя минеральных удобрений РМТ-4. а фосфорные в виде гранулированного суперфосфата - локально в гребни при посадке с помощью картофелепосадочнои машины КСМ-4. . Урожайность картофеля составила 388 ц/га, качество клубней было хорошее. В них содержалось сухих веществ 272. в пересчете на абсолютно сухое вещество. -. крахмал 22,3 азот 1.35 фосфор 0,53 калий...
Способ выбора этиопатогенетически обоснованного вида оперативного вмешательства при гастродуоденальной язве

Номер патента: 9261
Опубликовано: 30.06.2007
Автор: Соломонова Галина Алексеевна
МПК: A61B 1/273, A61B 5/05
Метки: этиопатогенетически, способ, оперативного, язве, обоснованного, гастродуоденальной, выбора, вмешательства, вида
Текст:
...в теле желудка,7 - электрод -зонда, расположенный в антральном отделе. Пример 1. Больной К., 52 лет. Диагноз хроническая язва двенадцатиперстной кишки, осложненная кровотечением, компенсированным стенозом. При эндоскопическом исследовании устанавливают наличие в двенадцатиперстной кишке язвы 1,2 см с тромбированным сосудом, деформацию кишки. Больному выполняют эндоскопический гемостаз. Через 10 суток с момента остановки...
Способ интегрирования генов в определенные участки генома клеток млекопитающих посредством гомологичной рекомбинации и векторы для его выполнения

Номер патента: 7288
Опубликовано: 30.09.2005
Авторы: РЕФФ, Митчелл, И., БАРНЕТТ, Ричард, Спенс, МакЛЭХЛЕН, Карен, Ретта
МПК: C12N 15/12, C12N 15/62, C12N 15/13...
Метки: участки, генома, векторы, посредством, рекомбинации, клеток, определенные, гомологичной, способ, генов, выполнения, интегрирования, млекопитающих
Текст:
...проистекают из явления гомологичной рекомбинации (при произвольной интеграции не будет продуцирован функциональный неоген и, соответственно, он не выживет в процессе селекции в присутствии 418). Таким образом,настоящее изобретение облегчает отбор для желаемой гомологичной процедуры. Более того, частота произвольных дополнительных интеграций в клетках, прошедших процесс гомологичной рекомбинации, оказывается низкой. Основываясь на упомянутом,...
Промотор синтазы ацетоксикислот для экспрессии генов в растении

Номер патента: 4799
Опубликовано: 30.12.2002
Авторы: ДИТРИХ, Габриэль, СМИТ, Джейн, ПЕНГ, Джианьинг
МПК: A01H 5/00, C12N 15/82, C12N 15/60...
Метки: генов, растении, экспрессии, промотор, ацетоксикислот, синтазы
Текст:
...радиоактивно помеченных РНК-зондов. Срезы тканей были препарированы и гибридизированы с РНК-зондами, кодирующими либосмысловую полосу (-), либоантисмысловую полосу . Для сравнения в качестве зондов использовались(малая субъединица ) смысловая полоса (-) илиантисмысловая полоса . В каждом случае ожидалось, что только антисмысловая полоса будет вступать в гибридизацию с мРНК, присутствующей в ткани.)зонд ) -зонд с)зонд ) -зонд. Фиг. 7 -...