Генно-инженерная конструкция, экспрессирующая кДНК гена CYP11А1 животного происхождения в растениях, и метод получения трансгенных растений с повышенной урожайностью и устойчивостью к фитопатогенам
Номер патента: 9201
Опубликовано: 30.04.2007
Авторы: Шпаковский Георгий Вячеславович, Манешина Татьяна Владимировна, Ярмолинский Дмитрий Георгиевич, Бердичевец Ирина Николаевна, Спивак Светлана Григорьевна, Бричкова Галина Геннадьевна, Картель Николай Александрович







Текст
(51)12 15/00 НАЦИОНАЛЬНЫЙ ЦЕНТР ИНТЕЛЛЕКТУАЛЬНОЙ СОБСТВЕННОСТИ ГЕННО-ИНЖЕНЕРНАЯ КОНСТРУКЦИЯ, ЭКСПРЕССИРУЮЩАЯ кДНК ГЕНА 11 А 1 ЖИВОТНОГО ПРОИСХОЖДЕНИЯ В РАСТЕНИЯХ, И МЕТОД ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ С ПОВЫШЕННОЙ УРОЖАЙНОСТЬЮ И УСТОЙЧИВОСТЬЮ К ФИТОПАТОГЕНАМ(71) Заявители Государственное научное учреждение Институт генетики и цитологии Национальной академии наук БеларусиГосударственное научное учреждение Институт биоорганической химии Национальной академии наук БеларусиИнститут биоорганической химии имени академиков М.М.Шемякина и Ю.А.Овчинникова Российской академии наук(72) Авторы Картель Николай Александрович Спивак Светлана Григорьевна Шпаковский Георгий Вячеславович Манешина Татьяна Владимировна Бричкова Галина Геннадьевна Ярмолинский Дмитрий Георгиевич Бердичевец Ирина Николаевна(73) Патентообладатели Государственное научное учреждение Институт генетики и цитологии Национальной академии наук БеларусиГосударственное научное учреждение Институт биоорганической химии Национальной академии наук БеларусиИнститут биоорганической химии имени академиков М.М.Шемякина и Ю.А.Овчинникова Российской академии наук(57) 1. Рекомбинантная плазмида 450 для экспрессии гена 111 в растениях, содержащая в составефрагмента длиной 2535 п.о. окаймленную последовательностями 35 промотора и терминатора вируса мозаики цветной капусты кДНК гена 111 длиной 1754 п.о., кодирующую белок-предшественник цитохрома Р 450 из коры надпочечника быка с лидерным -концевым экстрапептидом из 39 аминокислотных остатков с молекулярной массой 60,3 кДа, ген устойчивости к фосфинотрицину - генпод контролем -промотора, ген устойчивости к канамицину, участок начала репликациииз плазмидыширокого круга хозяев и последовательность полилинкера. 2. Способ получения трансгенных растений, содержащих кДНК гена 111, заключающийся в кокультивировании выделенных из листьев растений предназначенных для трансформации протопластов с рекомбинантной плазмидой 450 по п. 1, предварительно линеаризованной путем гидролиза эндонуклеазой рестрикции 1201 по уникальному сайту 5-/, культивировании трансформированных протопластов до образования колоний, переносе колоний на питательную среду с селективным агентом для каллусообразования и регенерации растений. 9201 1 2007.04.30 Изобретение относится к области генетической инженерии, конкретно - к созданию генно-инженерной конструкции (рекомбинантной плазмиды 450), способной экспрессировать кДНК гена 111 животного происхождения в растениях, а также к способу получения трансгенных растений, несущих цитохромы Р 450 животных и обладающих повышенной продуктивностью и устойчивостью к грибным фитопатогенам. В растениях обнаружено и изучено около 150 цитохромов, относящихся к классу цитохромов Р 450. Данные белки вовлечены во многие процессы первичного и вторичного метаболизма, включая биосинтез лигнинов, терпеноидов, стеринов, жирных кислот, гормонов, пигментов, фитоалексинов и т.д. Растительные цитохромы Р 450 участвуют как в регуляции процессов роста и развития, так и в индукции защитной реакции растений 1. Например, цитохром 90 является ключевым ферментом биосинтеза брассиностероидов (биологически активных стероидов растений) в арабидопсисе 2. Интрон-экзонная структура и последовательность консервативных и функционально-важных доменов 90 имеет высокую степень гомологии с таковыми Р 450-монооксигеназ млекопитающих, включая стероидные гидроксилазы, одной из которых является цитохром 450(4501) 3. Дефект (мутация) гена , кодирующего 90, приводит к нарушению роста гипокотиля у растений арабидопсиса. Это может быть обусловлено отсутствием С 23-гидроксилированных производных брассинолида катастерона, экдизонподобного растительного стероидного гормона 4. В последнее время для создания трансгенных растений, характеризующихся интенсивным ростом, развитием и повышенной устойчивостью к фитопатогенам используются природные растительные гены, контролирующие уровень таких гормонов роста, как, например, индолил-3-уксусная кислота 5, гены, кодирующие растительные липооксигеназы 6, а также гены других ферментов, катализирующих синтез важных регуляторных молекул. Известны наиболее близкие к заявленным ряд генно-инженерных конструкций, экспрессирующих различные варианты кДНК гена 7110, кодирующего один из цитохромов Р 450 сои (71), и способ получения трансгенных растений, включающий трансформацию генома табака указанными плазмидами 7. Однако получаемые в этом случае трансгенные растения, содержащие кДНК гена цитохрома 71, демонстрируют только устойчивость к узкому спектру гербицидов типа фенилмочевины и не приобретают других положительных с точки зрения биотехнологии качеств. Изобретение решает задачу создания генно-инженерной конструкции (рекомбинантной плазмиды 450), содержащей кДНК гена 111 из коры надпочечников быка,а также разработку способа получения трансгенных растений, экспрессирующих кДНК гена 111 животных, с улучшенной продуктивностью и устойчивостью к грибным фитопатогенам. Поставленная задача решается за счет того, что рекомбинантная плазмидная ДНК 450 содержит в составе -фрагмента длиной 2535 п.о. окаймленную последовательностями 35 промотора и терминатора вируса мозаики цветной капусты комплементарную ДНК гена 111 длиной 1754 п.о., кодирующий белок-предшественник цитохрома 450 из коры надпочечников быка с лидерным -концевым экстрапептидом из 39 аминокислотных остатков и суммарной молекулярной массой 60,3 кДа, и состоит из находящегося под контролем -промотора гена фосфинотрицин-ацетилтрансферазы (ген), обусловливающего устойчивость к фосфинотрицину полилинкера гена устойчивости к канамицину участка начала репликациииз плазмиды широкого круга хозяевпоследовательностейи , а также за счет того, что в способе получения трансгенных растений путем кокультивации их протопластов с рекомбинантной плазмидной ДНК, кокультивацию проводят с использованием рекомбинтной плазмидной ДНК 450, линеаризованной путем гидролиза эндонуклеазой рестрикции 120 по уникальному сайту 5-/. 2 9201 1 2007.04.30 Известно, что в коре надпочечников быка экспрессируется белок 4501, который катализирует начальную лимитирующую стадию стероидогенеза - отщепление боковой цепи холестерина и превращение его в прегненолон 8. Для осуществления заявленного изобретения из коры надпочечников быка вначале выделяют суммарную РНК, а затем из нее - фракцию мРНК. Используя полученный препарат мРНК в качестве матрицы, с помощью обратной транскрипции на ней синтезируют двухцепочечные кДНК, которые далее используют для создания кДНК клонотеки. При помощи гибридизации с радиоактивно меченным зондом, сконструированным на основе известной аминокислотной последовательности цитохрома 4501, отбирают клоны,несущие последовательность гена 111 из коры надпочечников быка. Фрагмент,представляющий собой полноразмерную кДНК незрелого 4501 и содержащий собственный инициирующий кодон, клонируют в векторе 51 под контролем 35 промотора. Из полученного промежуточного вектора с помощью эндонуклеаз рестрикции вырезают кассету, содержащую 35 промотор, кДНК гена цитохрома 4501 (450) и терминатор. Данную кассету встраивают в вектор 0229. Отбирают генно-инженерные конструкции, содержащие кассету в ориентации голова к хвосту (450) и хвост к хвосту (450) по отношению к генупод контролем -промотора, который присутствует в векторе 0229 изначально. Для трансформации растений табака методом кокультивации протопластов используют ДНК векторов, линеаризованную с помощью эндонуклеаз рестрикции. Отбор трансформантов проводят на селективных средах. Наличие встроенных генов определяют методами полимеразной цепной реакции (ПЦР) и гибридизации по Саузерну. Устойчивость растений к фитопатогенам определяют стандартными методиками. Последовательность выделенной нами кДНК гена 111 из коры надпочечников быка (1754 п.о.), использованной в дальнейшем для создания рекомбинантной плазмиды для ее экспрессии в растениях, приведена на фиг. 1. Карта полученной таким образом генно-инженерной конструкции, плазмиды 450,использованной в дальнейшем для трансформации растений, показана на фиг. 2. Использование плазмиды 450, описанной в данном изобретении, позволяет достаточно простым путем вводить кДНК гена 111 из коры надпочечников быка в геном растений для создания линий и сортов растений с более интенсивным ростом и с повышенной устойчивостью к фитопатогенам, чем исходная нетрансформированная форма. Изобретение иллюстрируют следующие примеры. Пример 1. Клонирование полноразмерной кДНК гена 111 из коры надпочечников быка. Тотальную клеточную РНК выделяют из коры надпочечников быка гуанидинтиоцианатным методом 9. 20 г ткани замораживают в жидком азоте и растирают до порошкообразного состояния, проводят экстракцию 40 мл лизирующего буфера следующего состава 50 мМ трис-гидрохлорид, рН 7,6, 25 мМ ЭДТА (этилендиаминотетрауксусная кислота), 0,07 мМ 2-меркаптоэтанол, 2-лаурилсаркозил. Полученный лизат трижды экстрагируют смесью фенол/хлороформ/изоамиловый спирт (2002004) до исчезновения интерфазы. Фазы разделяют центрифугированием 5 мин 5000 об/мин. Из водной фазы РНК осаждают 3 М ацетатом натрия (рН 6,05) в течение ночи в ледяной бане. Полученный осадок собирают центрифугированием, растворяют в ТЕ-буфере (0,05 мМ трис-гидрохлорид, рН 7,5, 0,01 мМ ЭДТА) и осаждают этанолом в присутствии 0,3 М хлористого натрия. Качество полученного препарата РНК исследуют методом гель-электрофореза в агарозном геле в присутствии формальдегида. Пробы растворяют в 20 мкл денатурирующего буфера следующего состава 3 мкл 36 формальдегида, 7 мкл формамида концентрированного и 8 мкл буфера для электрофореза (фосфоацетатный буфер). РНК прогревают в 3 9201 1 2007.04.30 течение 5 мин при 65 С и вносят в лунки 0,8 агарозного геля, приготовленного на электродном буфере, содержащем 1,1 М формальдегид. Гель окрашивают бромистым этидием(0,5 мкг/мл) и фотографируют в ультрафиолете. Препарат поли(А)-РНК получают хроматографией на олиго-целлюлозе 10. Спиртовой осадок РНК растворяют в 10 мл 0,01 М трис-гидрохлорида (рН 7,5), содержащем 0,5 М хлорид калия и наносят на колонку с олиго-целлюлозой. Колонку промывают 10-ю объемами раствора 0,5 М хлорида калия. РНК элюируют раствором 0,01 М трис-гидрохлорида, рН 7,5. На выделенной поли(А)-РНК синтезируют двухцепочечную кДНК далее ее встраивают с помощью соответствущего полилинкера в ДНК плазмидного вектора 13 по-сайту. Отобранные на селективной среде белые колонии (рекомбинантные клоны) анализируют методом гибридизации на фильтрах с 32 Р-меченным олигонуклеотидным зондом, комплементарным как экстрапептиду, так и началу зрелого фермента 4501. Радиоактивную метку вводят в зонд методом никтрансляции, который включает две ферментативные реакции введение в ДНК с помощью ДНКазыслучайно распределенных однонитевых разрывов (ников) с дальнейшим их расширением и одновременной достройкой с помощью ДНК-полимеразы. . Комплементарный олигонуклеотид инкубируют в смеси следующего состава 0,05 М трис-гидрохлорида (рН 7,5), 5 мМ хлорида магния, 10 мМ 2-меркаптоэтанол, 50 мкг/мл бычьего сывороточного альбумина, ДНК(примерно 0,1 мкг), все дезоксинуклеозидтрифосфаты (, , , ), в присутствии ДНКазыи ДНК-полимеразы 1 Е.(15, 30 мин). Меченый зонд выделяют из реакционной смеси на колонке с сефадексом -50. Соответствие клонированной последовательности кДНК гена 111 подтверждают посредством частичного секвенирования и рестрикционного анализа, а также гибридизацией с геномной ДНК быка по Саузерну 11. Полученная кДНК кодирует белокпредшественник 4501, состоящий из 520 аминокислотных остатков (а.о.), на 2 конце которого содержится экстрапетид (39 а.о.), обусловливающий способность белка проникать через митоходриальную мембрану, где он подвергается процессингу и приобретает каталитическую активность. Плазмида, несущая кДНК гена белка 4501, названа 13-450.- фрагмент кДНК 111 из 13-450 переклонируют по сайту в плазмидный вектор 1. Плазмиды 13-450 и 1 обрабатывают эндонуклеазой рестрикциив условиях, рекомендованных фирмой-производителем(Сибэнзим, Новосибирск) при четырехкратном избытке фермента. Реакцию проводят в объеме 25-50 мкл 2 ч при 37 С. Инкубационную смесь далее экстрагируют смесью фенол/хлороформ (11) рН 8,0, ДНК осаждают двумя объемами этилового спирта в присутствии 0,1 М хлорида натрия в ледяной бане и собирают центрифугированием в течение 10 мин при 5000 об/мин. Осадок растворяют в однократном трис-боратном буфере, фрагменты рестрикции разделяют при помощи агарозного гель-электрофореза в этом же буфере 12. Разделенные фрагменты ДНК выделяют из геля посредством электроэлюции. Электрофоретические зоны, содержащие нужные фрагменты, вырезают из геля, помещают в диализные мешки и подвергают электроэлюции в однократном трис-боратном буфере при напряжении 80 в течение 1 ч. Элюат экстрагируют смесью фенол/хлороформ (11) и очищают на микроколонке с сефадексом -50 в ТЕ-буфере, осаждают этиловым спиртом и проверяют гель-электрофорезом в агарозном геле. Легирование проводят при стандартных условиях 12 ч при 15 С при пятикратном избытке вставки по отношению к векторному фрагменту 12. Полученной лигазной смесью трансформируют клетки Е.109. Трасформацию проводят с применением 2 12. Из устойчивых к ампициллину рекомбинантных клонов выделяют плазмидную ДНК по методу Бирнбойма и Долли 13. Трансформированные клетки Е.109 выращивают в жидкой средес ампи 4 9201 1 2007.04.30 циллином (100 мкг/мл), содержащей 5 г(о.с.ч.), 8 г триптона (, США), 5 г дрожжевого экстракта (, США) в течение ночи при 37 С. Клетки (10 мл культуры) осаждают центрифугированием 3000 об/мин 20 мин, ресуспендируют в 200 мкл буфера следующего состава 25 мМ трис-гидрохлорида рН 8,0, 50 мМ глюкозы, 20 мМ ЭДТА,2 мг/мл лизоцима (, США), затем добавляют 200 мкл раствора 0,2 с 1, инкубируют до полного просветления суспензии в ледяной бане. Добавляют 150 мкл 3 М ацетата натрия рН 4,8, перемешивают и удаляют осадок центрифугированием(20 мин, 3000 об/мин). ДНК из супернатанта осаждают двумя объемами этилового спирта,осадок растворяют в 0,3 М ацетате натрия (рН 6,0) и осаждают двумя объемами этилового спирта. Осадок, собранный центрифугированием, растворяют в ТЕ-буфере. Физическое картирование полученной рекомбинантной ДНК проводят с помощью эндонуклеаз рестрикции , ,, . Пример 2. Получение плазмидной ДНК (плазмиды) 450. Удобным вектором для молекулярно-генетических манипуляций является плазмида 51. Она является производной плазмиды 18 и содержит в своем составе промотор и терминатор вируса мозаики цветной капусты 14. Между промотором и терминатором находятся сайты узнавания некоторых широко используемых рестриктаз ( , , , ). Удобным является также и то, что экспрессирующую кассету можно вырезать из вектора с помощью сразу нескольких рестриктаз (, , ,). Вектор является многокопийным и несет ген устойчивости к ампициллину. Полученная конструкция 450 является удобной при создании векторов, предназначенных для переноса гена 111 в геном растений, т.к. в ее последовательности удобно расположены сайты узнавания эндонуклеаз рестрикции, что позволяет легко вырезать кассету с геном 111 для переклонирования в других плазмидах. Для создания вектора 450 ДНК плазмиды 51 гидролизуют эндонуклеазой рестрициипри 30 С в буфере, содержащем 33 мМ трис-ацетат (рН 7,9 при 37 С), 10 мМ ацетат магния, 66 мМ ацетат калия, 0,1 мг/мл бычьего сывороточного альбумина. Затем после тепловой инактивации рестриктазы ДНК вектора дефосфорилируют инкубацией с 1 единицей щелочной фосфатазы при 37 С. Продукты реакций разделяют в 1 агарозном геле, содержащем 0,5 мкг/мл бромистого этидия. Фрагмент геля, содержащий линеаризованный вектор, вырезают с помощью скальпеля, затем из него с использованием процедуры замораживания-оттаивания выделяют ДНК. После очистки путем экстракции смесью фенол-хлороформ ДНК вектора легируют с -фрагментом, содержащим полноразмерную кДНК гена цитохрома 450 быка. Реакцию легирования проводят в течение ночи при 16 С с помощью ДНК лигазы фага Т 4. Легированную ДНК используют для трансформации штамма 5 (генотип -1 17 96 1 1 44 1 8015 8 -) по стандартной методике с использованием хлорида кальция (пример 1). Трансформированные клетки отбирают на агаризованной среде , содержащей 100 мкг/мл ампициллина. Для проверки наличия плазмиды отдельные колонии отсевают в пробирку с 4 млс ампициллином. Плазмидную ДНК выделяют, как было описано выше. Полученные препараты анализируют с помощью эндонуклеаз рестриции. Отбирают плазмиду, содержащую вставку последовательности,которая кодирует цитохром 450 быка. Проверяют правильность ориентации вставки с помощью рестрикции. При конструировании вектора для переноса последовательности кДНК гена цитохрома 450 быка в растения была использована плазмида 0229, которая представляет собой удобный вектор для трансформации растений 15. Она содержит под контролем-промотора ген фосфинотрицин-ацетил-трансферазы (ген ), который обусловливает устойчивость к фосфинотрицину. В векторе также имеется удобный для клонирования полилинкер, ген устойчивости к канамицину для отбора в бактериях, участок начала реп 5 9201 1 2007.04.30 ликациииз плазмиды широкого круга хозяев , что позволяет поддерживать плазмиду не только в, но и в. Гени поли линкер фланкированы участкамии , необходимыми для агробактериальной трансформации. Плазмиды 0229 и 450 гидролизуют энодонуклеазой рестрикциипри 37 С в буфере, содержащем 10 мМ трис- (рН 8,5), 10 мМ хлорид магния, 100 мМ хлорид калия, 0,1 мг/мл бычьего сывороточного альбумина. После тепловой инактивации рестриктазы ДНК вектора 0229 дефосфорилирилируют с помощью 1 единицы щелочной фосфатазы в той же реакционной смеси. Очистку фрагментов в агарозном геле,легирование и трансформацию проводят так же, как было описано выше. Отбор трансформантов проводят на агаризованной среде , содержащей 50 мкг/мл канамицина. После выделения плазмидной ДНК по методу щелочного лизиса ее анализируют с помощью эндонуклеаз рестрикции. Отбирают клоны, содержащие плазмиды со вставками в ориентации голова к хвосту (450) и хвост к хвосту (450) по отношению к генупод контролем -промотора 12. Схема полученной конструкции представлена на фиг. 2. Пример 3. Трансформация растений с использованием плазмидной ДНК 450 и анализ включения кДНК гена 111 в геном трансформированных растений и ее экспрессии в них. Подготовку ДНК плазмид 0229 (пустой вектор) и 450 (содержит нужный ген) для трансформации проводят следующим образом. ДНК выделяют из штамма Е.5 450 по методике, указанной в примере 1, в пересчете на больший объем. Для трансформации выделенную и очищенную плазмиду вначале линеаризуют с помощью соответствующей эндонуклеазы рестрикции. Вектор 450, который содержит последовательность гена 111 под контролем 35 промотора вируса мозаики цветной капусты линеаризуют путем гидролиза эндонуклеазой рестрикции 120 при 37 С в буфере, содержащем 10 мМ трис- (рН 7,5 при 37 С), 10 мМ хлорид магния, 0,1 мг/мл бычьего сывороточного альбумина. Для плазмиды 0229 используют гидролиз рестриктазойпри 37 С в буфере, содержащем 10 мМ трис- (рН 8,5 при 37 С),10 мМ хлорид магния, 100 мМ хлорид калия, 0,1 мг/мл бычьего сывороточного альбумина. Обработанные плазмиды (2-3 мкл) разделяют в 0,8 агарозном геле для подтверждения полноты рестрикции. ДНК плазмид осаждают этанолом, промывают 75 этанолом и высушивают в стерильном потоке воздуха. Плазмиду растворяют в стерильной дистиллированной воде, концентрацию доводят до 1 мкг/мкл. Получение трансгенных растений проводят на растениях табака. Могут быть использованы и другие двудольные и однодольные растения. Для трансформации используют асептическую культуру линии табака.1. Табак культивируют при 24/18, 16/8-часовом фотопериоде,освещенности порядка 2000-3000 лк. Для трансформации используют листья 6-8-недельных растений табака, выросших в стерильных условиях. Листья нарезают в растворе фермента и оставляют для выделения протопластов на 16-18 ч при температуре 24 С в темноте. Выделение протопластов осуществляют в ферментативном растворе следующего состава 0,4 М среда К 3 (макросоли К 3, микросоли В 5, хелатирующий агент, витамины К 3, ксилоза, сахароза, 2,4 , НУК,БАП (рН 5,8), агароза), целлюлаза -10, мацерозим -10, рН 5,6. На следующий день смесь фермента с протопластами фильтруют через металлический сеточку диаметром 100 мкм. Остатки тканей дополнительно промывают средой К 4 (макросоли К 3, микросоли В 5, хелатирующий агент, витамины К 3, ксилоза, сахароза, 2,4 ,НУК, БАП, рН 5,8) для полного выделения оставшихся протопластов. На протопласты наслаивают среду 5 (, 2, . глюкоза, рН 5,8) и центрифугируют 10 мин при 80. Хорошие протопласты собирают из интерфазы и дважды отмывают от фермента в 6 9201 1 2007.04.30 среде 5, центрифугируя по 5 мин при 70. Проводят подсчет общего количества протопластов в 5-среде. Для этого 100 мкл протопластов растворяют в 900 мкл среды 5, отбирают 10 мкл суспензии и подсчитывают жизнеспособные протопласты в гематоцитометре. Протопласты инкубируют в термостате 30 мин при 24 С, центрифугируют в течение 5 мин при 70, полностью удалив при этом среду 5, и ресуспендируют в среде МММ(манитол, 2, , рН 5,6) для получения 1000000 протопластов в 0,7 мл раствора. К протопластам (1 млн. на вариант) добавляют линеаризованную плазмиду (10 мкг), тимусную ДНК-носитель (40 мкг) и полиэтиленгликоль (0,7 мл). Через 15-20 мин проводят отмывку в среде 5. После центрифугирования при 70 в течение 5 мин удаляют надосадочную жидкость, и протопласты ресуспендируют в 0,5 мл среды К 3. Протопласты запаивают в среду К 3, содержащую 0,6 агарозы ( ). Культивирование запаянных протопластов проводят при 24 С в течение 1 суток в темноте, затем - при 24/18 С, 16/8 часовом фотопериоде, освещенности порядка 500 лк (рассеянный свет) в течение 5 суток. Колонии из трансформированных протопластов прорастают через 5-6 недель при 24/18 С, 16/8-часовом фотопериоде, освещенности порядка 500 лк в среде - (макросоли А-, микросоли В 5, витамины К 3, НУК, БАП, сахароза, манитол, рН 5,6) с селективным агентом фосфинотрицином (РРТ) 20 мкг/мл. Культивирование колоний для каллусообразования и регенерации проводят при 24/18 С, 16/8-часовом фотопериоде и освещенности порядка 2000-3000 лк. Спустя 5-6 недель, когда колонии достигают размера 2-3 мм в диаметре, их переносят на среду - (макросоли , микросоли В 5, хелатирующие агенты, 2 БАП,НУК, рН 5,6) с 0,6 агаром для каллусообразования и регенерации растений. Эффективность трансформации конструкциями 450 и 0229 - 32 независимых растения на 1000000 протопластов. Проростки размером 3-4 см пересаживают на среду - для укоренения. Укоренившиеся растения высаживают в почву для дальнейшего молекулярно-генетического, морфологического и патофизиологического анализов. Первичный отбор растений проводят по результатам теста на устойчивость к фосфинотрицину. Растения, укоренившиеся в почве, обрабатывают гербицидом фосфинотрицином в концентрации 40 и 80 мг/л для проверки наличия экспрессии гена . Выделение ДНК из отобранных растений проводят следующим образом. Растительные ткани, замороженные или свежие, растирают в ступке с жидким азотом. Гомогенат заливают буфером(100 М 1 рН 8,5, 100 М , 50 МрН 8,0, 2) в соотношении 12. После размораживания добавляют протеиназу К (10 мг/мл) прямо в ступку - 50-100 мкг на 1 мл смеси, инкубируют при 65 С 1,5-2 ч. После этого добавляют равный объем смеси фенол-хлороформ (11), перемешивают и центрифугируют при 4,5 тыс. оборотов в течение 20-30 мин. Снимают верхнюю фазу и переносят в новую пробирку, добавляют равный объем хлороформа, встряхивают, центрифугируют 20-30 мин при 4,5 тыс. оборотов. Верхний слой переносят в маленькие стаканчики на 50 мл, добавляют изопропанол в соотношении 0,6-0,9 от образца. С помощью палочки накручивают ДНК до полного ее извлечения. Палочки с намотанной на них ДНК промывают 75 спиртом и сушат около 40 мин. Высушенную ДНК растворяют в 300 мкл стерильного ТЕбуфера (101, 1, рН 8,0). После растворения ДНК обрабатывают РНКазой в течение 45-60 мин при 37 С. Затем проводят стандартную очистку фенол-хлороформом. ДНК осаждают этанолом, промывают 80 этанолом. После того как ДНК будет высушена, ее растворяют в ТЕ-буфере. Концентрацию ДНК определяют с помощью агарозного мини-гель электрофореза и с помощью спектрометрии. Пример 4. Фенотипический анализ трансгенных растений. Анализ изменений в фенотипе -трансгенных клонов табака и трансгенных растений, содержащих одновременно гени кДНК гена 111, проводят путем оценки и 7 9201 1 2007.04.30 сравнения с контрольными растениями таких морфологических параметров, как скорость укоренения, высота растений, время начала цветения, продуктивность растений. Фенотипический анализ проводили независимыми наблюдателями в начале, середине и конце вегетационного периода. Результаты приведены в табл. 1 на фиг. 3 представлена фотография контрольных и трансгенных растений табака в полевых условиях. Цифрами обозначены 1 - контрольное растение табака (высота 15 см), 2 - трансгенное растение табака, содержащее кДНК гена 111 (высота 72 см). Таблица 1 Сравнительная оценка морфологических признаков трансгенных растений табака,выращенных в условиях открытого грунта Начало Скорость Начало Высота Площадь Количество каллусо- укоренения, цветения, растения на листовой коробочек образовадни дата момент пластинки, с семенами ния, дни цветения,см 2 на растесм ние, шт Растение Контрольное нетрансгенное Контрольное с пустым вектором Трансгенное с кДНК гена 111 Трансгенные растения, содержащие кДНК гена 111, характеризуются более быстрыми процессами укоренения, более ранними сроками цветения и образования плодов(эти процессы происходят на 2 недели раньше по сравнению с контрольными растениями), более высокой продуктивностью (превосходит параметры контрольных растений в 3 раза), большей площадью листовой пластинки (больше по сравнению с контролем в 23 раза). Пример 5. Биотесты на белковых экстрактах из трансгенных растений, несущих кДНК гена 111 из коры надпочечников быка. На устойчивость к неспецифическим патогенам пасленовых - грибкурастения проверяют методом подсчета количества проросших спор в капле сока из листьев. Биотесты проводят следующим образом. В лунки микрокюветы наносят по 20 мкл суспензии спор(105 спор/мл), 20 мкл неочищенных белковых экстрактов из контрольных (нетрансформированных и трансформированных пустым вектором) и трансгенных растений, содержащих исследуемый ген. Микрокюветы инкубируют в течение 24 часов при 24 С, после чего под микроскопом проводят подсчет количества проросших спор. Полученные результаты приведены в табл. 2. Таблица 2 Оценка фунгицидных свойств трансгенных растений табака Количество непроросших Количество проросших спор,спор,Контрольное нетрансгенное 104 906 Контрольное с пустым вектором 156 858 Трансгенное с кДНК гена 111 8612 147 Растение 9201 1 2007.04.30 Средние арифметические значения количества проросших спор для контрольных трансгенных (с пустым вектором) и нетрансгенных линий табаков, варьируют от 80 до 96 . Для трансгенных табаков, содержащих кДНК гена 111, количество проросших спор варьирует от 7 до 21 . Эти результаты свидетельствуют о подавлении роста патогенных грибов экстрактами белка, выделенного из трансгенных растений, содержащих исследуемый ген. Пример 6. Биотесты на устойчивость трансгенных растений, содержащих кДНК гена 11 А 1, к грибным фитопатогенам. Опыты проводят на изолированных листьях трансгенных табаков, содержащих кДНК гена 111. В качестве тест-культуры используют фитопатогенные грибы вида, вызывающие пятнистость и раннюю гниль. В провоцирующих условиях листья среднего возраста и одного размера помещают в чашки Петри на фильтровальную бумагу,увлажненную водой. По 10 мкл суспензии спор возбудителя, содержащей 2000 спор/мл,наносят в 6 точек на лист, после чего чашки с листьями герметично закрывают для создания повышенной влажности и выдерживают в климокамере при температуре 24 С и 16 часовом световом дне. В качестве контроля используют листья нетрансформированных растений и растений, содержащих пустой вектор. Результаты биотеста на устойчивость к, оцененные по количеству и размеру некротических пятен на 10 сутки после заражения, представлены на фиг. 4. Цифрами обозначены 1 - контрольное нетрансгенное растение (поражение - 7 баллов) 2 - контрольное трансгенное растение с пустым вектором (поражение - 7 баллов) 3-5 - независимые линии трансгенных растений, содержащих кДНК гена 111 (3 - поражение 1 балл 4 - поражение 0 баллов 5 - поражение 2 балла) а - поражение листа. У устойчивых трансгенных линий табака, содержащих кДНК гена 111, средняя площадь пораженных на листьях участков на 10-е сутки была в 12 раз меньше, чем у контрольных растений, или же области некроза на листьях вообще отсутствовали. Источники информации 1..,.-450//. Национальный центр интеллектуальной собственности. 220034, г. Минск, ул. Козлова, 20. 11
МПК / Метки
МПК: C12N 15/00
Метки: фитопатогенам, конструкция, урожайностью, гена, трансгенных, получения, метод, происхождения, устойчивостью, экспрессирующая, генно-инженерная, растениях, кднк, cyp11а1, животного, растений, повышенной
Код ссылки
<a href="https://by.patents.su/11-9201-genno-inzhenernaya-konstrukciya-ekspressiruyushhaya-kdnk-gena-cyp11a1-zhivotnogo-proishozhdeniya-v-rasteniyah-i-metod-polucheniya-transgennyh-rastenijj-s-povyshennojj-urozhajjnosty.html" rel="bookmark" title="База патентов Беларуси">Генно-инженерная конструкция, экспрессирующая кДНК гена CYP11А1 животного происхождения в растениях, и метод получения трансгенных растений с повышенной урожайностью и устойчивостью к фитопатогенам</a>
Способ получения трансгенных свиней
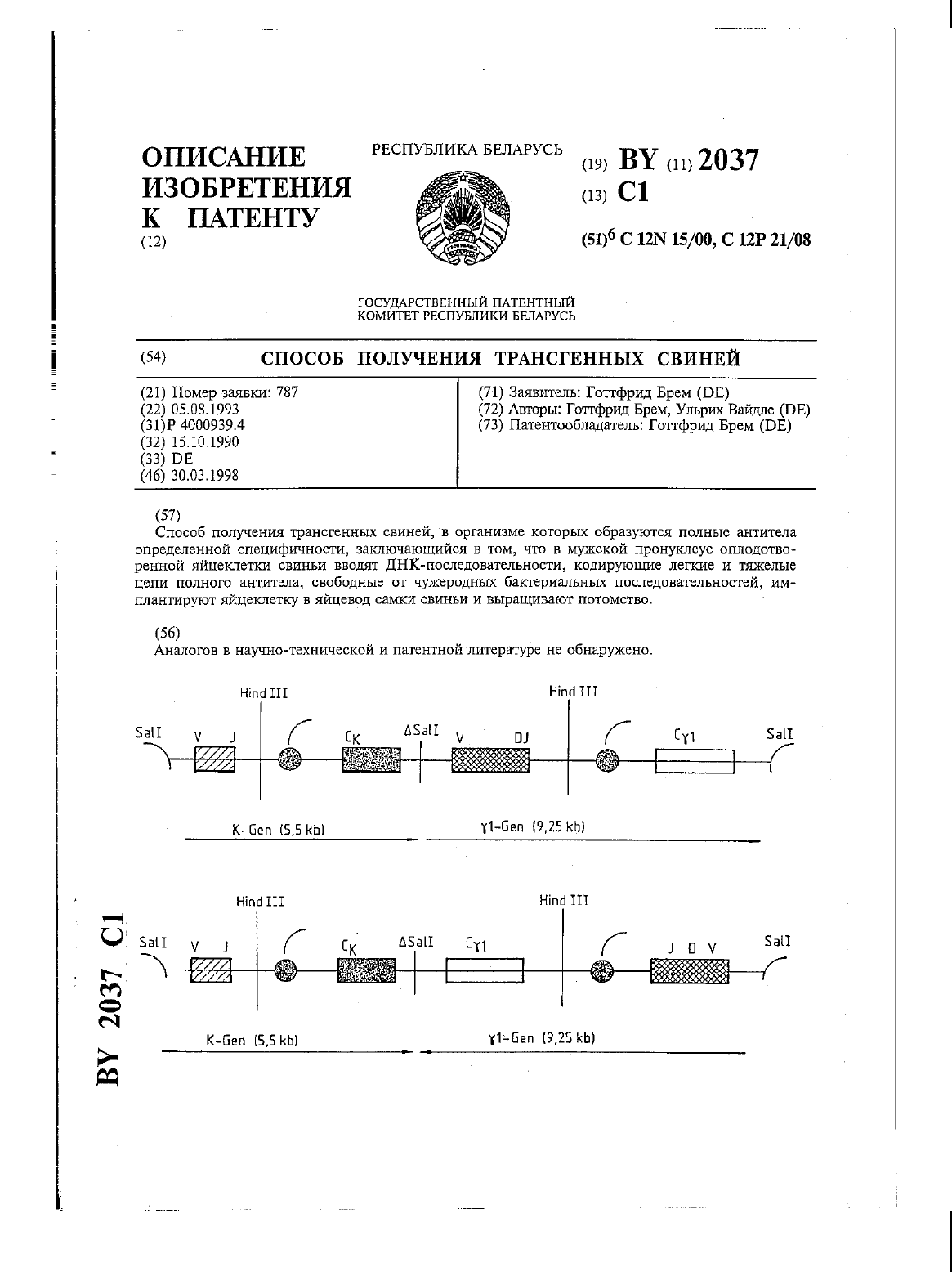
Номер патента: 2037
Опубликовано: 30.03.1998
Авторы: Ульрих Вайдле, Готтфрид Брем
МПК: C12P 21/08, C12N 15/00
Метки: получения, трансгенных, способ, свиней
Текст:
...часов после введения НСС животных осеменяют. Эмбрионы получают через 24-27 часов после оплодотворения. Для хирургического получения эмбрионов экивотных подвергают наркозу с помощью 160 мг азаперона (стресснила) и 400 мг метомидатгидрохлорида. После подготовки поля операции кожу открывают разрезом около 10 см длиной по срединной линии на высоте последних двух пар сосков и выкладывают матку, яйцевод и яичник. Матку перфорируют тупым...
Способ активации экспрессии транскрипционно “молчащего” гена, способ модификации экспрессии гена (варианты), эукариотическая клеточная линия (варианты), геном эукариотической клеточной линии

Номер патента: 3183
Опубликовано: 30.12.1999
Автор: СКОТТ С. ЧЕППЕЛЬ
МПК: C12N 1/11, C12N 15/11, C12N 15/67...
Метки: линии, клеточной, гена, модификации, геном, варианты, транскрипционно, эукариотической, линия, способ, эукариотическая, активации, клеточная, экспрессии, молчащего
Текст:
...регуляторной последовательности. Также в случае очень низкой эффективности гомологичной рекомбинации может появиться необходимость исключить амплификаторный ген, поскольку отношение негомологичной ДНК к гомологичной ДНК непосредственно связано с эффективностью гомологичной рекомбинации (, , 117759-770, 1987). Кроме того, возможно исключить ген положительной селекции и отобрать клетки только путем скринирования на продукцию желаемого белка...
Продуцирование трегалозы в растениях

Номер патента: 4187
Опубликовано: 30.12.2001
Авторы: Петер Йосеф Мария ВАН ДЕН ЭЛЬЗЕН, Мирьям Петронелла ДУС, Андреас ХУКЕМА, Ян ПЕН
МПК: A01N 3/00, A01H 5/00, A23L 3/3562...
Метки: растениях, продуцирование, трегалозы
Текст:
...сообщает устойчивость к гербицидам, полученным на основе глютатиона (ЕР-А 256 223), ген глютаминсинтетазы, который, определяя суперэкспрессию этого фермента, придает устойчивость к ингибиторам глютаминсинтетазы, таким, как фосфинотрицин ( 87/05327), ген ацетилтрансферазы из, который придает устойчивость к селективному агенту фосфинотритрицину (ЕР-А 275 957), ген, кодирующий 5-энолшикимат-3-фосфат-синтазу, придающий толерантность к...
Защитно-стимулирующий комплекс для защиты семян и растений от болезней и регулирования роста растений (варианты), способ защиты семян и растений от болезней и регулирования роста растений
Номер патента: 8953
Опубликовано: 28.02.2007
Автор: Клименко Владимир Иванович
МПК: A01N 61/00, A01C 1/00, A01N 59/00...
Метки: роста, защиты, регулирования, болезней, способ, варианты, защитно-стимулирующий, комплекс, растений, семян
Текст:
...З, но из-за недостаточной стабильности составов после 6-ти месяцев хранения в составах образовывались крупные включения и наблюдались случаи забивания распылительных систем опрь 1 скивателей. Крупные включения в известных составах 6 и 7 снижали их фунгицидную эффективность в результате недостаточной степени эффективности покрытия поверхности растений рабочей жидкостью. Защитно-стимулирующий комплекс Полиазофос К и способ его применения 6...
Способ получения твердого препарата для защиты культурных растений и твердый препарат для защиты культурных растений

Номер патента: 3240
Опубликовано: 30.03.2000
Авторы: УЭДЛОК, Дейвид Джон, ДЕ ЛИНД ВАН ВЕЙНГАРДЕН, Герхард
МПК: A01N 25/14, A01N 25/08, A01N 25/10...
Метки: твердый, растений, защиты, способ, твердого, препарат, культурных, препарата, получения
Текст:
...Каждому специалисту ясно, что оптимизация рабочей температуры и соотношений компонентов может быть осуществлена для каждого активного ингредиента путем рутинного экспериментирования. В соответствии с этим может быть использован активный ингредиент, имеющий температуру плавления в диапазоне от 60 до 200 . Конкретными активными ингредиентами, подходящими для использования в способе,согласно настоящему изобретению, являются...