Способ повышения терморезистентности стандартной молоди осетровых рыб
Номер патента: 13383
Опубликовано: 30.06.2010
Авторы: Шалак Михаил Владимирович, Барулин Николай Валерьевич, Плавский Виталий Юльянович




Текст
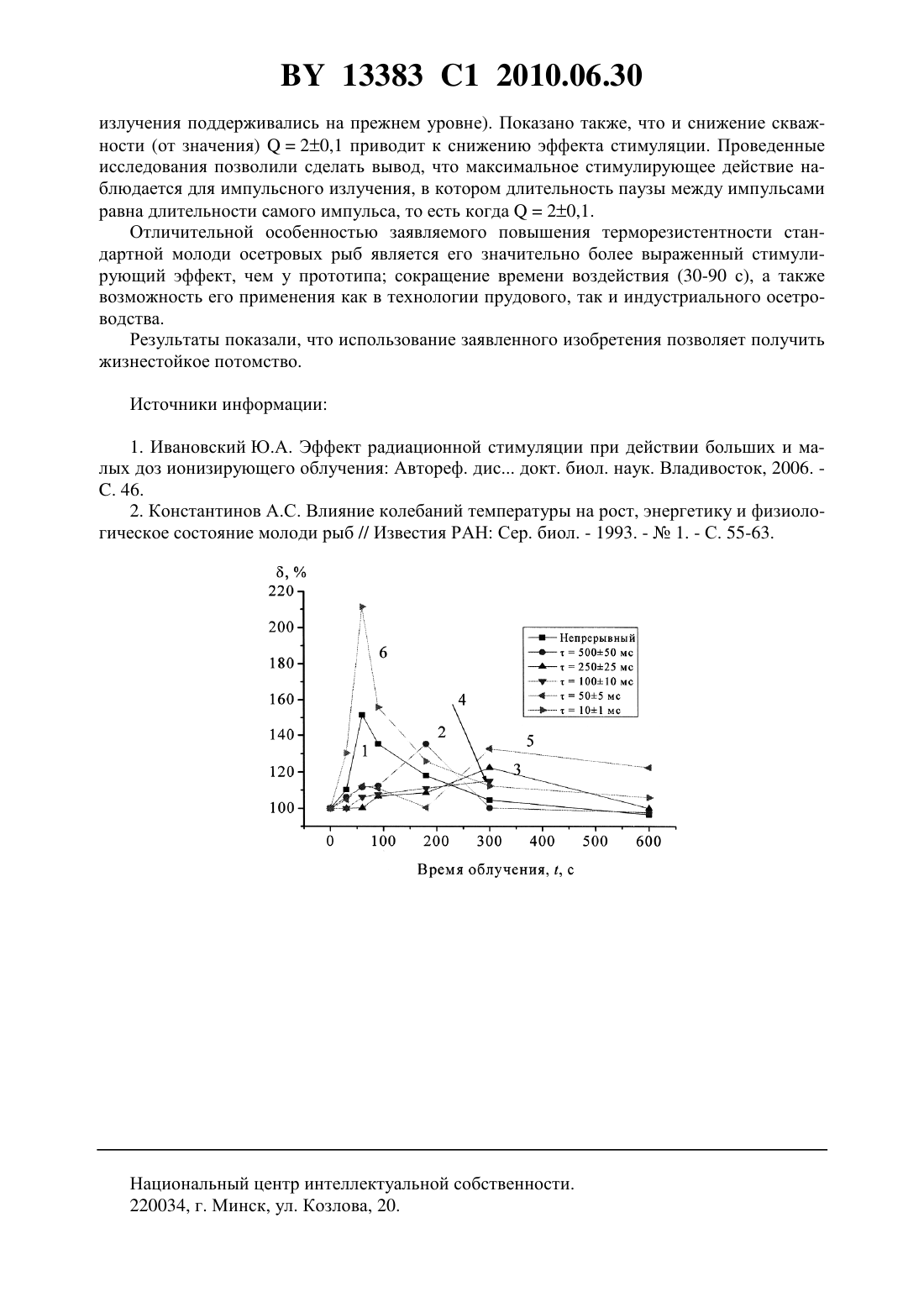
(51) МПК (2009) НАЦИОНАЛЬНЫЙ ЦЕНТР ИНТЕЛЛЕКТУАЛЬНОЙ СОБСТВЕННОСТИ СПОСОБ ПОВЫШЕНИЯ ТЕРМОРЕЗИСТЕНТНОСТИ СТАНДАРТНОЙ МОЛОДИ ОСЕТРОВЫХ РЫБ(71) Заявители Государственное научное учреждение Институт физики имени Б.И.Степанова Национальной академии наук Беларуси Учреждение образования Белорусская государственная орденов Октябрьской Революции и Трудового Красного Знамени сельскохозяйственная академия(72) Авторы Барулин Николай Валерьевич Шалак Михаил Владимирович Плавский Виталий Юльянович(73) Патентообладатели Государственное научное учреждение Институт физики имени Б.И.Степанова Национальной академии наук Беларуси Учреждение образования Белорусская государственная орденов Октябрьской Революции и Трудового Красного Знамени сельскохозяйственная академия(57) Способ повышения терморезистентности стандартной молоди осетровых рыб, заключающийся в том, что на эмбрионы осетровых рыб на стадии органогенеза воздействуют в течение 30-90 с импульсным поляризованным лазерным излучением ближней инфракрасной области спектра с длиной волны 85020 нм, длительностью импульса 101 мс,скважностью 20,1 при средней плотности мощности 2,70,2 мВт/см 2. Изобретение относится к рыбоводству, а именно к способам выращивания молоди осетровых рыб, и может найти широкое применение как в технологии прудового, так и индустриального осетроводства. Известен способ повышения терморезистентности гидробионтов (жаброногий рак науплии), включающий облучение яицгамма-радиацией в дозировке 10 Гр 1. Согласно 1, при повышении температуры воды - среды обитания половозрелых рачков - до 36 С в течение 1 суток наблюдается гибель всех необлученных особей (контрольная группа). Рачки, предварительно облученные в дозе 10 Гр (опытная группа), показали значительно большую терморезистентность, чем контрольные интактные гидробионты. При этом в опытной группе летальный исход через 1 сутки отмечен у 63 особей. При дозе 20 Гр гибельпри их содержании при температуре 36 С была несколько выше, чем в контрольной группе. Недостатком известного способа повышения терморезистентности гидробионтов является потенциальное побочное действие радиоактивного излучения в отношении обслу 13383 1 2010.06.30 живающего персонала, а также опасность накопления радионуклидов в товарной продукции (мясе). По этой причине известный способ не применим в технологии прудового и индустриального осетроводства. Наиболее близким к заявляемому способу является способ повышения терморезистентности осетровых рыб к экстремальным температурам, основанный на выращивании рыб (с момента перехода личинок на внешнее питание), в условиях переменного терморежима среды обитания (прототип) 1. Переменный терморежим осуществляют в форме колебания температуры воды с различным периодом и амплитудой. Контролем служат рыбы, содержащиеся при среднем значении альтерирующих температур, а также их верхнем и нижнем значениях. Согласно данным 1, в условиях переменного терморежима, когда верхнее значение колеблющихся температур не выходят за пределы физиологической нормы, отмечается ускорение роста рыб по сравнению с режимом постоянного поддержания температуры на заданном уровне. Наиболее оптимальным режимом для выращивания осетра является следующий 1 раз в сутки температура воды в течение 2 ч повышается на 3-4 С. Затем за такой же промежуток времени вода, постепенно охлаждаясь, возвращается к исходному значению. Для осетровых рыб максимальный эффект наблюдается при варьировании температуры в диапазоне 234 С. При указанных условиях наблюдается ускорение роста молоди, а также повышается верхнее значение пороговой экстремальной температуры (п), приемлемое для указанного вида рыб. Так, если при поддержании постоянной температуры на уровне 19 С или 23 С, п 31 С то при 234 С,пор 32,5 С. При этом в процессе выращивания молоди в условиях колебательного терморежима наблюдается снижение (по сравнению со стационарным терморежимом) отхода(гибели) молоди. Недостатком известного способа является недостоверное отличие в пороговых экстремальных температурах, характерных для колебательного терморежима по сравнению со стационарным. Кроме того, данный способ технически сложен для осуществления в условиях индустриального осетроводства и неосуществим в условиях прудового рыбоводства. Задачей заявленного изобретения является получение жизнестойкого потомства, повышение эффективности воспроизводства и выращивания молоди осетровых рыб, а также оптимизация технологии товарной аквакультуры. Поставленная задача решается в способе повышения терморезистентности стандартной молоди осетровых рыб, заключающийся в том, что на эмбрионы рыб на стадии органогенеза воздействуют в течение 30-90 с импульсным поляризованным лазерным излучением ближней инфракрасной области спектра с длиной волны 85020 нм длительностью импульса 101 мс, скважностью 20,1 при средней плотности мощности Р 2,70,2 мВт/см 2. Сущность предлагаемого способа заключается в следующем. Увлажненную оплодотворенную икру возвратного гибрида бестера (стерлядьбестер (1 на стадии органогенеза подвергают воздействию поляризованного лазерного излучения. Облучение осуществляют с помощью полупроводникового лазера. Излучение лазера расфокусируют линзой таким образом, чтобы размер светового пятна соответствовал размеру монослоя облучаемой икры. Технические характеристики полупроводникового лазера обеспечивают возможность воздействия лазерным излучением в непрерывном и импульсном режимах при длительности импульса 101 505 10010 25025 и 50050 мс. Средняя плотность мощности (в мВт/см 2) лазерного излучения, воздействующего на эмбрионы, определяют по формуле Р/, где- измеренная средняя мощность лазерного излучения в мВт- площадь светового пятна (в см 2) на уровне монослоя икры. При воздействии лазерным излучением в непрерывном и импульсном режимах средняя мощность излучения составляет 2705 мВт. Выравнивание средней мощности излучения для указанных режимов воздействия осуществляют путем регулировки тока,2 13383 1 2010.06.30 протекающего через полупроводниковый лазер, и контролируют измерителем средней мощности и энергии лазерного излучения ИМО-3 С. Для определения оптимального времени воздействия, оказывающего максимальный стимулирующий эффект на терморезистентность стандартной молоди осетровых рыб к облучению икры проводят в течение 30 60 90 180 300 600 с. Полученные зависимости стимулирующего действия лазерного излучения (, ) от времени воздействия для различных режимов воздействия представлены на фигуре. На фигуре по оси абсцисс отложено время облучения в с, а по оси ординат - величина стимулирующего действия лазерного излучения(Тр / Трк)100 , где Трк - продолжительность (мин) выживания молоди осетровых рыб при температуре 32 С, эмбрионы которой не подвергались воздействию лазерного излучения, то есть терморезистентность для контрольной группы Тр продолжительность (мин) выживания молоди осетровых рыб при температуре 32 С, эмбрионы которой на стадии органогенеза подвергались воздействию поляризованным лазерным излучением инфракрасной области спектра, то есть терморезистентность для опытной группы. Причем кривая 1 на фиг. 1 соответствует варианту воздействия на эмбрионы на стадии органогенеза непрерывным инфракрасным лазерным излучением 85020 нм средней плотностью мощности Р 2,70,2 мВт/см 2. Кривая 2 получена при облучении эмбрионов импульсным инфракрасным лазерным излучением 85020 нм средней плотностью мощности Р 2,70,2 мВт/см 2 при длительности импульса 500 мс кривая 3 - при длительности импульса 250 мс кривая 4 - при длительности импульса 100 мс кривая 5 - при длительности импульса 50 мс кривая 6 - при длительности импульса 10 мс. Контрольные (интактные) образцы икры выдерживают в тех же условиях, что и опытные образцы. Количество икринок в опытной и контрольной группах составляет 300 шт. количество повторностей - 3. Статистическую обработку полученных данных проводят по стандартной методике с использованием пакета. Достоверность полученных данных оценивают по критерию Стьюдента. Температура икры в момент ее облучения лазером (а также в контрольных образцах) составляет 161 С. Икру после лазерного облучения помещают в уменьшенные образцы аппаратов Вейса,предназначенные для инкубирования оплодотворенной икры, где, достигнув завершения эмбрионального развития, происходил выклев предличинок. Каждые опытные и контрольные группы содержат в отдельных аппаратах Вейса. В процессе инкубирования икры обеспечивается постоянство гидрохимических условий. Выклюнувшихся предличинок переносят в отдельные садки для каждой исследуемой группы. Плотность предличинок составляет 5000 шт/м 2. Переход на активное питание происходит через 101 суток. При этом плотность посадки личинок сокращают в 2 раза и она составляет 2500 шт/м 2. В первые 10 дней после перехода на активное питание кормление осуществлялось науплиями артемии и стартовыми комбикормами в соотношении 6040 в последующем - 100 сухими комбикормами. Кормление осуществляют каждые 2 часа. В период выращивания температура воды составляет 202 С. Контроль над гидрохимическим режимом осуществляют ежедневно. Достигнув 50 суток, стандартного для рыбопосадочного материала осетровых рыб, выращенных в заводских условиях, проводят тесты на терморезистентность. Опыты на определение терморезистентности проводят в 80 литровом аквариуме, на котором были смонтированы нагреватель мощностью два киловатта, фильтр - насос с микропроцессором для перемешивания и принудительной аэрации воды, реле для автоматического поддержания заданной температуры, контактный термометр и садочек из латунной сетки, в который помещались подопытные личинки. Опыты ставят по следующей схеме. Аквариум заполняют водой, в которой выдерживают в течение 24 ч молодь без пищи. Затем включают нагреватель, и в течение 40-50 мин подогревают воду до 32 С. Время терморезистентности отсчитывают с момента достижения 32 С. Содержание ки 3 13383 1 2010.06.30 слорода в воде аквариума поддерживают на уровне не ниже 7 мг/л. Гибель молоди определяли по остановке движения жаберных крышек. В момент гибели каждой особи фиксируют время терморезистентности. Установлено, что воздействие на эмбрионы осетровых рыб на стадии органогенеза поляризованным лазерным излучением инфракрасной области спектра с длиной волны 85020 нм плотностью мощности Р 2,70,2 мВт/см 2 способно оказывать стимулирующее действие на терморезистентность стандартной заводской молоди осетровых рыб. В таблице приведены значения времени (в мин) выживания молоди осетровых рыб с момента достижения экстремальной температуры (32 С), эмбрионы которой на стадии органогенеза не подвергались (контрольная группа) и подвергались (опытная группа) воздействию поляризованного лазерного излучения инфракрасной области спектра. Причем данные, приведенные в таблице для импульсного излучения, получены при скважностиТ/20,1, где Т - период световых колебаний в мс,-длительность импульса лазерного излучения в мс. То есть, при скважности 2 длительность светового импульса равна длительности паузы (п) между импульсами, поскольку Тп. Время выживания молоди осетровых с момента достижения экстремальной температуры (32 С), эмбрионы которой на стадии органогенеза не подвергались(контрольная группа) и подвергались (опытная группа) воздействию поляризованного лазерного излучения инфракрасной области спектра с длиной волны 85020 нм, плотностью мощности Р 2,70,2 мВт/см 2 в непрерывном Режим воздействия, длиТерморези- Величина стиму- Достоверность Время обтельность импульса,стентность, лирующего дейотличий от лучения, с мин ствия,контроля, мс Контроль 0 100 145,31,1 непрерывный 60 220,01,6 151,450,6 Р 0,001 180 импульсный,500 мс 196,52,9 135,32,0 Р 0,001 300 импульсный,250 мс 178,24,1 122,42,3 Р 0,001 600 импульсный,100 мс 167,82,3 115,51,5 Р 0,001 300 импульсный,50 мс 132,62,0 192,74,0 Р 0,001 30 импульсный,10 мс 189,31,5 130,283,1 Р 0,001 60 импульсный,10 мс 307,03,5 211,32,4 Р 0,001 90 импульсный,10 мс 226,42,6 155,81,8 Р 0,001 180 импульсный,10 мс 182,96,5 125,94,5 Р 0,001 Из представленных данных следует, что облучение эмбрионов осетровых рыб на стадии органогенеза приводит к значительному увеличению терморезистентности, при которой наблюдается увеличение времени выживания молоди при нахождении их в экстремальной температуры (32 С). Так, если в контрольной (необлученной) группе продолжительность терморезистентности составила Трк 145,31,1 мин, то для группы рыб,эмбрионы которой подвергались воздействию в течение 60 с импульсного лазерного излучения с длиной волны 85020 нм, плотностью мощности Р 2,70,2 мВт/см 2 с длительностью импульса 101, Тр 307,03,5 мин (достоверность отличий от контроля Р 0,001). То есть гибель молоди, эмбрионы которой подвергалась воздействию оптического излучения, наблюдается при более продолжительном воздействии на нее экстремальной температуры, чем у необлученных особей. Отметим, что при воздействии непрерывного излучения с теми же параметрами (85020 нм плотность мощности Р 2,70,2 мВт/см 2) максимальное отличие от контроля наблюдается при 60 с. В дан 4 13383 1 2010.06.30 ном варианте воздействия Тр 220,01,6 мин (достоверность отличий от контроля Р 0,001). Таким образом, представленные данные свидетельствуют, что воздействие поляризованным лазерным излучением как в непрерывном, так и в импульсном режимах на эмбрионы осетровых рыб на стадии органогенеза обеспечивает повышение терморезистентности стандартной молоди осетровых рыб. Величина стимулирующего эффекта терморезистентности зависит от времени и режима воздействия лазерным излучением. На фигуре приведены зависимости величины стимулирующего эффектаот времени воздействия на эмбрионы осетровых рыб на стадии органогенеза непрерывного лазерного излучения, а также импульсного лазерного излучения, характеризующегося длительностью импульса 101 505 10010 25025 и 50050 мс. Как следует из фигуры, кривая 6, максимальное повышение терморезистентности наблюдается при воздействии импульсным лазерным излучением в течение 60 с 101 мс и составляет 211,32,4 . Увеличение или уменьшение времени воздействия при 101 мс приводит к снижению стимулирующего эффекта. Так, при времени воздействия 30 с стимулирующий эффект составляет 130,283,1 , а при 90 с 155,81,83 . Из данных, показанных на фигуре, кривая 6 следует, что дальнейшее увеличение времени облучения эмбрионов при 101 мс приводит к значительному снижению эффекта стимуляции. Таким образом, при облучении эмбрионов осетровых рыб на стадии органогенеза поляризованным лазерным излучением инфракрасной области спектра с длиной волны 85020 нм плотностью мощности Р 2,70,2 мВт/см 2 максимальная стимуляция термоустойчивости молоди осетровых рыб наблюдается для импульсного лазерного излучения с 101 мс при времени воздействия 30-90 с. Стимуляция терморезистентности стандартной молоди осетровых рыб наблюдается также и при воздействии непрерывным и импульсным лазерным излучением(85020 нм, Р 2,70,2 мВт/см 2) с длительностью импульсов 505 10010 25025 и 50050 мс. Однако, как следует из фигуры во всех остальных вариантах импульсного воздействия 50050 мс (кривая 2)25025 мс (кривая 3)10010 мс (кривая 4)505 мс (кривая 5) а также непрерывного воздействия (кривая 1) стимулирующий эффект значительно ниже, чем при 101 мс. Обращает на себя внимание, что оптимальное время воздействия для стимуляции устойчивости к дефициту кислорода зависит от длительности импульсов. Так, если для непрерывного излучения максимальный эффект стимуляции наблюдается для 60 с, то для импульсного излучения при 50050 мс для 180 с при 25025 мс 300 с 10010 мс - для 600 с а при 505 мс 300 с. Таким образом, максимальное стимулирующее действие в отношении терморезистентности при облучении оплодотворенной икры на стадии органогенеза поляризованным лазерным излучением инфракрасной области спектра с длиной волны 85020 нм наблюдается для импульсного режима (101 мс) воздействия плотностью мощности Р 2,70,2 мВт/см 2 в течение 30-90 с. Установлено, что важным параметром, определяющим эффективность стимулирующего действия лазерного излучения, выражающегося в повышении терморезистентности стандартной молоди осетровых рыб, является не только длительность импульса лазерного излучения, но и продолжительность темнового периода (паузы) между импульсами (п), то есть скважность. Например, если при длительности импульса 10 мс, скважности 20,1, Р 2,70,2 мВт/см 2, длительности воздействия 60 с величина стимулирующего действия составляет 211,32,4 , то при увеличении скважности до 30,1 величина стимулирующий эффект составляет 142,32,8(остальные характеристики 5 13383 1 2010.06.30 излучения поддерживались на прежнем уровне). Показано также, что и снижение скважности (от значения)20,1 приводит к снижению эффекта стимуляции. Проведенные исследования позволили сделать вывод, что максимальное стимулирующее действие наблюдается для импульсного излучения, в котором длительность паузы между импульсами равна длительности самого импульса, то есть когда 20,1. Отличительной особенностью заявляемого повышения терморезистентности стандартной молоди осетровых рыб является его значительно более выраженный стимулирующий эффект, чем у прототипа сокращение времени воздействия (30-90 с), а также возможность его применения как в технологии прудового, так и индустриального осетроводства. Результаты показали, что использование заявленного изобретения позволяет получить жизнестойкое потомство. Источники информации 1. Ивановский Ю.А. Эффект радиационной стимуляции при действии больших и малых доз ионизирующего облучения Автореф. дис докт. биол. наук. Владивосток, 2006. . 46. 2. Константинов А.С. Влияние колебаний температуры на рост, энергетику и физиологическое состояние молоди рыб // Известия РАН Сер. биол. - 1993. -1. - С. 55-63. Национальный центр интеллектуальной собственности. 220034, г. Минск, ул. Козлова, 20. 6
МПК / Метки
МПК: A01K 61/00
Метки: стандартной, терморезистентности, молоди, способ, рыб, осетровых, повышения
Код ссылки
<a href="https://by.patents.su/6-13383-sposob-povysheniya-termorezistentnosti-standartnojj-molodi-osetrovyh-ryb.html" rel="bookmark" title="База патентов Беларуси">Способ повышения терморезистентности стандартной молоди осетровых рыб</a>
Способ повышения токсикоустойчивости стандартной молоди осетровых рыб

Номер патента: 12356
Опубликовано: 30.08.2009
Авторы: Плавский Виталий Юльянович, Барулин Николай Валерьевич, Шалак Михаил Владимирович
МПК: A01K 61/00
Метки: токсикоустойчивости, молоди, способ, рыб, стандартной, осетровых, повышения
Текст:
...Температура икры в момент ее облучения лазером (а также в контрольных образцах) составляла 161 С. Икру после лазерного облучения помещают в уменьшенные образцы аппаратов Вейса,предназначенные для инкубирования оплодотворенной икры, где, достигнув завершения эмбрионального развития, происходил выклев предличинок. Каждые опытные и контрольные группы содержат в отдельных аппаратах Вейса. В процессе инкубирования икры обеспечивается...
Способ повышения устойчивости стандартной молоди осетровых рыб к дефициту кислорода

Номер патента: 12355
Опубликовано: 30.08.2009
Авторы: Плавский Виталий Юльянович, Барулин Николай Валерьевич, Шалак Михаил Владимирович
МПК: A01K 61/00
Метки: кислорода, устойчивости, стандартной, осетровых, дефициту, рыб, способ, повышения, молоди
Текст:
...образцы. Количество икринок в опытной и контрольной группах составляет 300 шт. количество повторностей - 3. Статистическую обработку полученных данных проводят по стандартной методике с использованием пакета. Достоверность полученных данных оценивают по критерию Стьюдента. Температура икры в момент ее облучения лазером (а также в контрольных образцах) составляла 161 С. Икру после лазерного облучения помещают в уменьшенные образцы аппаратов...
Способ стимуляции размерно-весовых показателей молоди осетровых рыб

Номер патента: 12358
Опубликовано: 30.08.2009
Авторы: Плавский Виталий Юльянович, Барулин Николай Валерьевич, Лашкевич Анатолий Иванович
МПК: A01K 61/00
Метки: осетровых, стимуляции, способ, рыб, молоди, размерно-весовых, показателей
Текст:
...икру помещают для дальнейшего инкубирования в уменьшенные образцы аппаратов Вейса, где, достигнув завершения эмбрионального развития, происходит выклев предличинок. Опытные и контрольные группы содержат в отдельных аппаратах, в которых обеспечивают постоянство гидрохимических условий. Выклюнувшихся предличинок с каждой группы переносят в отдельные пронумерованные пластиковые садки, входящие в общую систему водоснабжения осетрового...
Способ выращивания обеззараженных личинок и молоди рыб в системе оборотного водоснабжения

Номер патента: 3864
Опубликовано: 30.06.2001
Авторы: Иващенко Сергей Алексеевич, Макаревич Викентий Викентьевич, Иващенко Василий Алексеевич, Иващенко Алексей Иванович
МПК: A01K 61/00
Метки: оборотного, системе, обеззараженных, водоснабжения, выращивания, личинок, молоди, способ, рыб
Текст:
...воды в системе по предлагаемому способу до 175 кг, в установке пос. Рыбное до 21 кг. Причем расход электроэнергии на 1 кг рыбопосадочного материала составляет 8 кВт/ч,что в 3 раза ниже, чем при известном способе, а расход воды снижается почти в 5 раз. Использование озона для подготовки воды в системе оборотного водоснабжения позволяет в одной технологической операции одновременно производить очистку воды от органо-минеральных загрязнений,...
Способ повышения локальной концентрации кислорода в биологических тканях пациента

Номер патента: 9855
Опубликовано: 30.10.2007
Авторы: Асимов Мустафо Мухамедович, Рубинов Анатолий Николаевич, Асимов Рустам Мустафович
МПК: A61N 5/067
Метки: концентрации, кислорода, повышения, локальной, биологических, тканях, способ, пациента
Текст:
...функции к бактериям. Величина ТсРО 2 (транскутаного напряжения О 2), непосредственно указывает на содержание О 2 в коже и его состояния метаболизма. Этот параметр является определяющим 2 9855 1 2007.10.30 для установления эффективности заживления ран, особенно после ампутации конечности,когда наступает хроническая ишемия. Экспериментально установлено, что лечение ран существенно усложняется в условиях тканевой гипоксии, т.е....
Предыдущий патент: Способ диагностики перенесенной транзиторной ишемической атаки в каротидном бассейне
Следующий патент: Отключаемый колесный узел транспортного средства
Случайный патент: Тормозная рычажная передача тележки рельсового транспортного средства